

 Курсовая работа: Загальна характеристика і особливості життєдіяльності бактерій родини Enterobacteriaceae
Курсовая работа: Загальна характеристика і особливості життєдіяльності бактерій родини Enterobacteriaceae
Більшість ентеробактерій можуть використовувати в якості субстратів для дихального метаболізму велику кількість простих органічних сполук – органічні кислоти, амінокислоти, вуглеводи. В аеробних умовах, всі ці бактерії добре ростуть на звичайних складних бактеріологічних середовищах, азотисті компоненти яких (амінокислоти і пептиди) забезпечують їх окислюючими субстратами. В анаеробних умовах, однак, ріст стає строго залежним від наявності зброджує мого вуглеводу. Деякі моносахариди, дисахариди і поліспирти зброджуються всіма представниками даної групи. Використання полісахаридів менш розповсюджено.
Хоча ентеробактерії вирощують на складних середовищах, мінімальні харчові потреби цих організмів, зазвичай прості. Представники багатьох родів взагалі не потребують факторів росту (наприклад Escherichia coli, багато видів Salmonella), а у ауксотрофних організмів ці потреби зазвичай прості. Особливо розповсюджена потреба в нікотиновій кислоті, що характерна для багатьох видів роду Proteus. У Salmonella typhi є специфічна потреба в триптофані.
Дослідження механізму регуляції біосинтезу амінокислот у багатьох ентеробактерій виявило характерні особливості цього процесу, які відрізняють данні організми від всіх інших бактерій. Наприклад, перша стадія в біосинтезі амінокислот родини аспартату, перетворення аспарагінової кислоти в аспартилфосфат, у ентеробактерій завжди здійснюється трьома ізофункціональними аспартаткіназами: аспартаткіназою I, яка і інгібується, і репресується треоніном, аспартаткіназою II, яка репресується, але не інгібується метіоніном, і аспартаткіназою III, яка і інгібується, і репресується лізином. Цей тип регуляції аспарагінового шляху не був виявлений ні в однієї бактерії не кишкової групи.
ГЦ-склад ДНК ентеробактерій варіює в широких межах – від 37 до 63%. За виключенням роду Proteus, представники якого суттєво відрізняються за нуклеотидним складом. ГЦ-склад ДНК споріднених організмів трьох видів - Salmonella, Escherichia і Shigella – розрізняються зовсім мало. Діапазон змінення
ГЦ-складу для „класичних” бактерій родини Enterobacteriaceae (від 37 до 59%) дуже близький до діапазону для представників родини з полярним джгутикуванням (від 39 до 63%) [10].
Бродіння
Зброджування сахаридів у ентеробактерій відбувається за шляхом Ембдена-Мейєргофа. Продукти бродіння у різних представників групи розрізняються і якісно, і кількісно. Однак у цих процесів є одна характерна біохімічна властивість, яка рідко зустрічається при інших типах бактеріального бродіння. Мова йде про особливий спосіб розщеплення однієї з проміжних сполук – піровиноградної кислоти (ПВК), що веде до утворення мурашиної кислоти:
СН3СОСООН + КоАSН→СН3СОSКоА+ НСООН.
Таким чином, мурашина кислота часто є основним кінцевим продуктом бродіння. Вона накопичується, хоча не завжди, оскільки деяки з цих бактерій володіють формнатгідрогенліазою, яка розщеплює мурашину кислоту до СО2 і Н2:
НСООН → СО2+Н2.
У таких організмів замість мурашиної кислоти як кінцевого продукту бродіння утворюються еквімолярні кількості Н2 і СО2.
Найбільш розповсюдженим типом перетворення сахаридів при бродінні у ентеробактерій є так зване бродіння змішаного типу, яке приводить до утворення молочної, оцтової, бурштинової і мурашиної кислот (або СО2 і Н2), а також етилового спирту. Такий тип бродіння властив представникам родів Salmonella, Escherichia, Shigella і Proteus. Відношення кількості різних кінцевих продуктів різниться як від штаму до штаму, так і для одного штаму при рості в різних умовах, наприклад при різних значеннях рН. Така різниця відображає ту обставину, що кінцеві продукти утворюються з піровиноградної кислоти трьома незалежними шляхами (рис. 3.1). Утворення газу в результаті зброджування сахаридів є дуже важливою ознакою для ентеробактерій – газоутворювальні бактерії роду Escherichia відрізняються за цим критерієм від патогенних видів групи Shigella і Salmonella typhi, які зброджують сахариди без виділення газу. В випадку простого бродіння змішаного типу газ може утворюватися тільки за рахунок розщеплення мурашиної кислоти; відповідно, утворення газу відображає наявність форміатгідрогенліази. Цей ферментний комплекс, звісно, не суттєвий для бродіння і може загубитися в результаті мутації без зміни здатності бактерії до бродіння. І дійсно, дослід показує, що в природі існують „неаерогенні”, тобто не утворюючі газоподібних продуктів штами такого типового продуценту їх, як Escherichia coli. Тому, хоча виділення газу являє собою корисну ознаку при класифікації ентеробактерій, даний критерій не є безпомилковим.
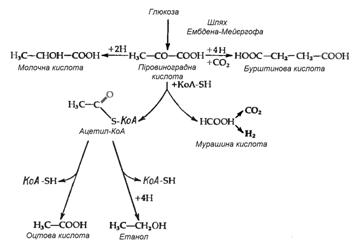
Рисунок 3.1 - Шляхи утворення із ПВК типових кінцевих продуктів кислотного бродіння змішаного типу (за [10])
Іншою ознакою, яка має велике значення для ідентифікації ентеробактерій, є здатність зброджувати дисахарид лактозу, що обумовлено наявністю у цих організмів β-галактозідази. Ефективність зброджування лактози залежить ще і від наявності специфічної пермеази галактозидів, яка сприяє проникненю лактози в клітину. Штами які мають β-галактозідазу, але позбавленні пермеази, не можуть поглинати лактозу зі швидкістю, яка достатня для інтенсивного бродіння, і зазвичай класифікуються як не здатні до зародження цього сахариду. Зброджування лактози властиво Escherichia, але відсутні у Salmonella, Shigella і Proteus. Деякі штами Shigella утворюють β-галактозідазу, але не можуть зброджувати лактозу із-за відсутності пермеази [10].
4. ГЕНЕТИКА БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Відкриття явища переносу генів у ентеробактерій при кон’югації і трансдукції дозволило доволі детально дослідити генетичні родинні відносини між деякими її представниками. Утворення хромосомних гібридів між E. coli і бактеріями родів Salmonella і Shigella каже про дуже високу ступінь генетичної гомології цих бактерій, що підтверджується і порівнянням хромосомних карт двох найбільш вивчених в цьому відношенні видів − Salmonella typhimurium і Escherichia coli. Як видно на рис. 4.1 і 4.2, де представленні в спрощеному вигляді хромосомні карти цих двох організмів, багато маркерів розташовуються на обох хромосомах в однакових локусах.
Про близьку генетичну спорідненість груп Escherichia, Salmonella і Shigella свідчить також висока ступень ДНК-ДНК-гібридизації in vitro. Утворення ж хромосомних гібридів між бактеріями цієї підгрупи і представниками інших родів ентеробактерій (Proteus, Enterobacter) відбувається дуже рідко. Крім того, випробування по гібридизації ДНК-ДНК вказують, що ступень генетичної гомології між представниками підгрупи Escherichia− Salmonella− Shigella та іншими групами ентеробактерій доволі низька. Ентеробактерії, що належать до різних родів, можуть отримувати плазміни від донорних штамів E. coli при кон’югації і потім зберігати їх як позахромосомні елементи (ефективність кон’югації варіює в широких межах; таблиця 4.1). Таким чином серед бактерій кишкової групи можуть розповсюджуватися F-фактори, які містять додаткові гени (наприклад F-lac), і R-фактори, які визначають стійкість до різноманітних лікарських засобів [10].
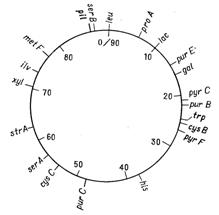
Рисунок 4.1 - Генетична карта E. coli К12 в спрощеному вигляді (за [10])
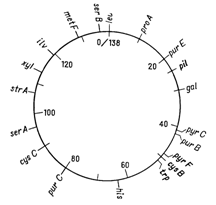
Рисунок 4.2 - Генетична карта S. typhimurium в спрощеному вигляді (за [10])
5. ЕКОЛОГІЯ БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Середовищем проживання для більшості ентеробактерій є кишковий тракт хребетних тварин і людини.
В організмі людини багато ентеробактерій містяться в складі мікробних біоценозів тонкого і товстого кишечника. Патогенні види зустрічаються тільки у хворих і бактеріоносіїв.
З випорожненнями людей і тварин ентеробактерії потрапляють в навколишнє середовище. В ній вони можуть зберігатися на протязі різних строків в залежності від виду і умов. Деякі з них (E. coli) використовуються в санітарній мікробіології в якості показників фекального забруднення навколишнього середовища [3].
Кишкова паличка наприклад може зберігатися в воді і ґрунті декілька місяців [11].
Широке розповсюдження сальмонел в природі і різноманіття шляхів, якими вони проникають в організм людини, пояснюється їх генетичною пластичністю. Сальмонели легко знаходять екологічні ніші і адаптуються до найрізноманітніших умов [12].
Клебсієли – широко розповсюджені бактерії. Вони порівняно стійкі до факторів навколишнього середовища завдяки наявності капсули і можуть на протязі тривалого часу зберігатися в ґрунті, воді, приміщеннях. Разом з тим вони зустрічаються в складі мікробних біоценозів в організмі людини і тварин [3].
Протеї P. vulgaris і P. mirabilis є мешканцями кишечника багатьох тварин, виявляються також в стічних водах і ґрунті. Всі види протеїв можуть виділятися з хворих людей [3].
6. АНТИГЕНИ БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Антигенна будова служить одним із суттєвих критеріїв, на яких основана класифікація, а також ідентифікація ентеробактерії. Розрізняють три основних типи антигенів: 1) О-соматичний антиген; 2) Н-джгутиковий антиген; 3)К-антигени. О-антиген є складовою частиною ліпополісахариду (ЛПС) зовнішнього шару клітинної стінки. Специфічність О-антигену визначається детермінантними сахаридами (гексозами і аміносахаридами), ковалентно зв’язаними з базисною частиною ЛПС. Н-антиген локалізується в джгутиках клітини. Він складається з білка флагеліну. Капсульні антигени К-антигени, так як і О-антигени, містяться в ЛПС клітинної стінки, але в більш поверхневому шарі. Вони маскують О-антигени.
В зв’язку з цим для визначення О-антигену у бактерій, які володіють К-антигеном, необхідно зруйнувати останній шляхом кип’ятіння або автоклавування культури. К-антигени за хімічними властивостями відносяться до кислих полісахаридів. До К-антигенів відносяться К-антигени E. coli, М-антигени Salmonella schottmuelleri, Vi-антиген збудника черевного тифу, який виявляється також у S. parathyphi C та деяких штамів E. coli. Всі ці антигени характеризуються імуннохімічною специфічністю, що дозволяє диференціювати роди і види, а також виділяти серед них серогрупи і серологічні варіанти (серовари). Антигенну будову ентеробактерій вивчають в реакціях аглютинації, непрямої (пасивної) гемаглютинації, преципітації, зокрема при імунно-електрофорезі та інших імунологічних реакціях з відповідними діагностичними сироватками.
Страницы: 1, 2, 3, 4, 5, 6, 7, 8
