

 Реферат: Симбиоз
Реферат: Симбиоз
 |
Однако все высказанные по этому поводу точки зрения до сих пор 'остаются лишь догадками и большей частью не подтверждены экспериментально: лишайники оказались очень трудным объектом для физиологических исследований. Ученые пока не научились выращивать и поддерживать в живом состоянии слоевище лишайников в искусственных условиях. Тот контакт между грибом и водорослью, который с такой легкостью достигается в природе (достаточно вспомнить многообразие лишайников!), никак не удается воспроизвести в лабораторных условиях. Наоборот, при переносе лишайников в лабораторию этот контакт легко нарушается и растение просто погибает. Время от времени появляются сообщения об удачных опытах выращивания лишайника в условиях лаборатории, но пока эти сообщения единичны и не всегда достоверны.
Одной из причин неудач подобных попыток можно считать чрезвычайно медленный рост лишайников. Лишайники — многолетние растения. Обычно возраст взрослых слоевищ, которые можно увидеть где-нибудь в лесу на стволе деревьев или на почве, составляет не менее 20—50 лет. В северных тундрах возраст некоторых кустистых лишайников рода кладония достигает 300 лет. Слоевище лишайников, имеющих вид корочки, в год дает прирост всего 0,2—0,3 мм.
Кустистые и листоватые лишайники растут несколько быстрее — в год их слоевище увеличивается на 2—3 мм. Поэтому, чтобы вырастить взрослый лишайник в лаборатории, требуется не менее 20 лет, а может быть, и вся жизнь исследователя. Трудно проводить столь долговременный эксперимент!
Вот почему физиологические особенности лишайников, в том числе взаимоотношения компонентов, как правило, изучают на культурах изолированных мико- и фикобионтов. Этот метод очень перспективен, так как позволяет ставить длительные и воспроизводимые опыты. Но, к сожалению, данные, полученные этим методом, не могут полностью отразить те процессы, которые происходят в целом слоевище лишайника.
И тем более мы не вправе считать, что в природе, в естественных условиях, в слоевищах лишайника эти процессы протекают точно так же, как в культурах изолированных симбионтов. Вот почему все теории, пытающиеся объяснить взаимоотношения компонентов лишайников, остаются пока лишь догадками.
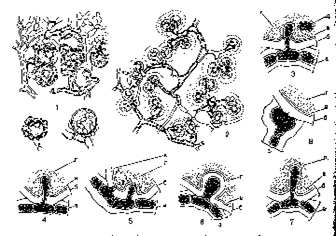
Более успешным оказалось изучение форм контакта между гифами гриба и клетками водорослей в слоевищах лишайников. Как показали исследования с применением электронной микроскопии, в слоевище лишайников можно встретить по крайней мере пять типов контакта между грибными гифами и водорослевыми клетками (рис. 289).
Чаще всего отдельная клетка водоросли и клетка грибной гифы находятся в непосредственном контакте друг с другом. В таком случае гриб образует специальные абсорбционные, всасывающие органы, которые проникают внутрь водорослевой клетки или плотно прижимаются к ее оболочке.
В настоящее время среди абсорбционных органов гриба в слоевище лишайников различают несколько типов: гаустории, импрессории и аппрессории.
Формы контакта между гифами гриба и клетками водорослей в слоевище лищайников.
Гаустории— это боковые выросты гиф гриба, которые прорывают оболочку клетки водоросли и проникают в ее протопласт (рис. 289, 2). Обычно в клетке водоросли развивается одни гаусторий, но иногда их может быть и два. В слоевище лишайника гаустории встречаются в большом количестве и существуют продолжительное время. Было замечено, что в оболочках молодых гаусториев нет отложений целлюлозы, которая могла бы затруднять обмен между клеткой водоросли и гифой гриба. Старые гаустории почти всегда одеты довольно толстым слоем целлюлозы. Различают интрацеллюлярные (внутриклеточные) и интрамембранные (внутриоболочковые) гаустории.
Интрацеллюлярные гаустории полностью прорывают оболочку клетки водоросли и проникают глубоко внутрь ее протопласта (рис.289, 3). Интрацеллюлярные гаустории образуются в случае резкого паразитизма гриба на водоросли. Это особенно характерно для лишайников с примитивным строением слоевища.
У более высокоорганизованных лишайников образуются только интрамембранные гаустории. Они прорывают оболочку клетки водоросли и достигают ее протопласта, но не углубляются в него, а остаются в оболочке водорослевой клетки (рис. 289, 5). Наибольшее количество интрамембранных гаусториев образуется в слоевище лишайников весной, в начале вегетационного периода. С наступлением осени они далеко отступают от протопласта водоросли.
Другой тип всасывающих органов гриба — импрессории— тоже боковые выросты грибных гиф, но, в отличие от гаусториев, они не прорывают оболочку клетки водоросли, а вдавливают ее внутрь (рис. 289, 6, 7). Импресcopии отмечены у очень многих лишайников, например у широко распространенной пельтигеры (Peltigera).
Интересно, что в слоевищах, произрастающих во влажных местообитаниях, импрессории почти не развиваются, у тех же видов в сухих местообитаниях они образуются в большом количестве. При длительной засухе число импрессориев также увеличивается. Предполагают, что в засушливые периоды и в сухих местообитаниях гриб, чтобы удовлетворить потребности в питании, увеличивает свою всасывающую поверхность за счет увеличения количества и размеров импрессориев.
В отличие от гаусториев и импрессориев, образованных боковыми отростками гифы, аппрессории образуются вершиной грибной гифы. Такая вершина гифы плотно прижимается снаружи к оболочке клетки водоросли, никогда не проникая ни в ее протопласт, ни в ее внутренний слой (рис. 289, 8).
Наличие в слоевищах многих лишайников абсорбционных органов гриба хорошо доказывает паразитическую сущность отношений микобионта к фикобионту. Но во многих случаях у лишайникового гриба все же не удается обнаружить особых абсорбционных органов, чаще всего у лишайников, фикобионт которых имеет тонкие оболочки клеток. В таких случаях уже внешний контакт гифы гриба и клетки водоросли может обеспечить обмен веществами между ними. Так, например, обстоит дело у многих видов рода кладония. Фикобионтом кладонии является одноклеточная зеленая водоросль требуксия. У этих лишайников отдельные клетки водорослей окружены со всех сторон тонкими тонкостенными гифами, иногда поделенными на мелкие клеточки. Эти гифы, которые носят название обволакивающих или контактных, не проникают в протопласт клеток водоросли и не внедряются в их оболочку, а просто окружают клетки со всех сторон, так что каждая из них становится похожей на маленький шар, охваченный пальцами рук (рис. 289,7). Иногда гифы полностью оплетают водоросли в виде сплошного покрова и при этом, сливаясь своими стенками, даже образуют клеточную псевдопаренхимную ткань. На первый взгляд кажется, что водоросли не особенно страдают от плотного окружения гифами гриба: они долго сохраняют свою зеленую окраску и продолжают интенсивно делиться.
Но в более старых участках слоевища можно найти немало отмерших обесцвеченных клеток — гриб рано или поздно все-таки убивает водоросли. Такой же тип контакта между гифами гриба и клетками водорослей был найден у некоторых слизистых и базидиальных лишайников.
У ряда лишайников, в слоевище которых встречаются нитчатые улотриксовые водоросли, можно наблюдать еще один тип контакта. Как правило, в таком случае нити водорослей бывают целиком покрыты грибными гифами. Причем лишь иногда гифы образуют на поверхности водорослевой нити рыхлую сетку. Чаще же они располагаются очень густо и, срастаясь своими стенками, образуют сплошной чехол. Отдельная лопасть такого лишайника имеет вид тончайшего волоса. Под микроскопом она напоминает полую трубку, стенки которой образованы сросшимися грибными гифами; внутри трубки тянется нить водоросли.
У слизистых лишайников семейства коллемовых (Collemataceae) обычно не наблюдается никакого контакта между грибными гифами и клетками водорослей. Слоевище коллемовых не имеет дифференцированной структуры: нити водоросли носток разбросаны в беспорядке среди грибных гиф по всей толще слоевища (рис. 297, 2). Никаких абсорбционных отростков в клетках водорослей обычно не заметно; гифы гриба и нити сине-зеленой водоросли расположены друг около друга, не вступая в видимый контакт. Предполагают, что в данном случае гриб поглощает органические вещества, ассимилируемые водорослями, прямо из слизи, которая обычно окружает нити ностока. Однако более тщательное изучение этих лишайников показало, что у многих видов коллемы (Collema) в слоевище время от времени образуются специальные абсорбционные гифы, которые тесно прижимаются к одной из клеток водорослевой нити, а через некоторое время можно наблюдать отмирание этой клетки.
Описанные выше формы контакта между гифами микобионта и клетками водорослей, по всей видимости, не исчерпывают всего многообразия способов, с помощью которых гриб и водоросль в слоевище лишайников устанавливают между собой тесную связь. Исследования в этом направлении только начинаются. Можно думать, что дальнейшее изучение тончайших структур лишайникового слоевища с помощью электронного микроскопа не только даст много нового в описании физических контактов между грибным и водорослевым компонентами лишайников, но и откроет новые горизонты в понимании их взаимоотношений
_________________________________________
Пока лишайники являют собой единственный строго доказанный случай возникновения одного совершенно нового организма из двух. Этот факт послужил толчком для поиска синтетических форм в самых различных систематических группах растений и животных. Однако все усилия, приложенные в данном направлении, оказались пока безрезультатными. Тем не менее, предположение о возможности существования синтетических форм организмов оказалось настолько заманчивым, что в биологии появляется новое течение. В отличие от обычных, прочно утвердившихся взглядов биологов на происхождение организмов как на переход от простого к сложному путем дифференциации, зарождается новое представление о возникновении сложного организма из более простых путем синтеза. Некоторые биологи стали рассматривать растительную клетку не как продукт постепенной дифференциации протопласта, а как симбиотический комплекс, синтезированный из нескольких простых организмов. Эти идеи зародились и получили наиболее полное развитие в трудах наших отечественных ученых.
Впервые предположение о важной роли формативного симбиоза (т.е. симбиоза, приводящего к образованию новых форм) в эволюции организмов было высказано академиком А. С. Фаминциным в 1907 г. Развивая дальше эти мысли, К. С. Мережковский в 1909 г. сформулировал гипотезу симбиогенного происхождения организмов и назвал ее “теорией симбиогенезиса”. В дальнейшем она получила широкую известность среди биологов. В 20-е годы ее поддержал и развил дальше известный советский ботаник Б. М. Коз о-П о ля нс кий. В наши дни эти идеи, уже на новом уровне развития биологии, были возрождены американской исследовательницей Сага н-М а р гу л и с в ее гипотезе происхождения эукариотических клеток. В соответствии с этой гипотезой такие клеточные органеллы, как митохондрии, базальные тела жгутиков и пластиды эукариотических клеток, возникли из симбиотирующих в них прокариотических клеток сине-зеленых водорослей и бактерий. В качестве основного довода приводятся некоторые черты сходства в составе, строении и поведении перечисленных органелл и прокариотов. Бесспорно, эти факты заслуживают самого пристального внимания. Однако они недостаточны для обоснования гипотезы симбиогенеза, поскольку черты сходства, как известно, могут появляться у разных по происхождению структур или организмов II вследствие параллелизма в эволюции. Так, система стигма — жгутик у золотистых, желто-зеленых и бурых водорослей по внешнему виду и функциям очень напоминает палочки сетчатки глаза животных, хотя весь процесс их заложения и ход онтогенетического развития говорят о том, что об общности происхождения этих образований не может быть и речи.
Исследование симбиотических организмов в электронном микроскопе показывает, что даже у такого высокоразвитого симбиоза, как глаукоцистис, партнеры сохраняют свои индивидуальные черты и автономность. Анализ симбиозов водорослей с различными организмами выявляет определенную направленность в развитии отношений между партнерами, главным образом по линии максимальной специализации функций и вызванных этим обстоятельством структурных перестроек при сохранении их как самостоятельных организмов; это идет вразрез с положениями гипотезы симбиогенеза. Все это свидетельствует о том, что в настоящее время гипотеза симбиогенеза находится на той стадии разработки, когда логические построения явно преобладают над фактами.
Безусловно, симбиоз может привести к созданию новых организмов, что подтверждается появлением такой своеобразной растительной группы, как лишайники. Отрицать роль симбиоза в эволюции нельзя. И все же очевидно, что это не единственный и не основной путь образования новых форм жизни. С одной стороны, к такому выводу подводит факт существования именно лишайников, поскольку они образуют крайне специализированную и обособленную группу организмов, представляющих слепую ветвь филогенетического развития. С другой стороны, сейчас накапливается большой фактический материал по тонкой организации клетки. Он дает возможность воссоздать картину вероятного обособления и усложнения организации некоторых клеточных органелл у водорослей. Кстати, именно отсутствие подобного рода фактов в свое время как раз и стимулировало зарождение гипотезы симбиогенеза.
Список использованной литературы: “Жизнь растений” (Том 3; Стр72 “Сожительство водорослей с другими организмами”.-Т. В. Седова; Стр385 “Взаимоотношения гриба и водоросли в теле лишайника” Н. С. Голубкова.)
Под ред. профессора М. М. Голлербаха.
Москва “Просвещение” 1977 год
487 стр.
Лекции Н. К Христофоровой “Биотические отношения” (Симбиоз)
