

 Реферат: Роль материнского генома в развитии потомка
Реферат: Роль материнского генома в развитии потомка
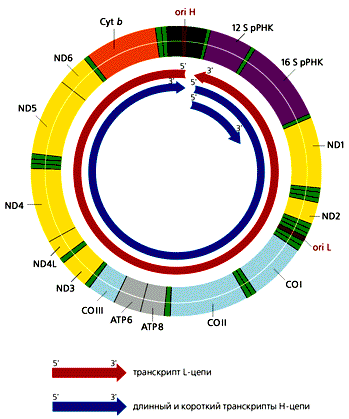 В митохондриях общее число молекул с
D-петлей значительно превыша-ет число полностью реплицирующихся молекул.
Обусловлено это тем, что у D-петли есть дополнительные функции — прикрепление
мтДНК к внутренней ме-мбране и инициация транскрипции, поскольку в этом районе
локализованы промоторы транскрипции обеих цепей ДНК. В отличие от большинства
эв-кариотических генов, которые транскрибируются независимо друг от друга, ка-ждая
из цепей мтДНК млекопитающих переписывается с образованием одной молекулы РНК,
начинающейся в районе ori H. Помимо этих двух длинных мо-лекул РНК,
комплементарных Н- и L-цепям, формируются и более короткие участки Н-цепи, которые
начинаются в той же точке и заканчиваются на 3'-кон-це гена 16S рРНК (рис.4).
Таких коротких транскриптов в 10 раз больше, чем длинных. В результате
созревания (процессинга) из них образуются 12S рРНК и 16S рРНК, участвующие в
формировании митохондриальных рибосом, а так-же фенилаланиновая и валиновая
тРНК. Из длинных транскриптов вырезаются остальные тРНК и образуются
транслируемые мРНК, к 3'-концам которых при-соединяются полиадениловые
последовательности. 5'-концы этих мРНК не кэ-пируются, что необычно для
эвкариот. Сплайсинга (сращивания) не происхо-дит, поскольку ни один из
митохондриальных генов млекопитающих не содер-жит интронов.
В митохондриях общее число молекул с
D-петлей значительно превыша-ет число полностью реплицирующихся молекул.
Обусловлено это тем, что у D-петли есть дополнительные функции — прикрепление
мтДНК к внутренней ме-мбране и инициация транскрипции, поскольку в этом районе
локализованы промоторы транскрипции обеих цепей ДНК. В отличие от большинства
эв-кариотических генов, которые транскрибируются независимо друг от друга, ка-ждая
из цепей мтДНК млекопитающих переписывается с образованием одной молекулы РНК,
начинающейся в районе ori H. Помимо этих двух длинных мо-лекул РНК,
комплементарных Н- и L-цепям, формируются и более короткие участки Н-цепи, которые
начинаются в той же точке и заканчиваются на 3'-кон-це гена 16S рРНК (рис.4).
Таких коротких транскриптов в 10 раз больше, чем длинных. В результате
созревания (процессинга) из них образуются 12S рРНК и 16S рРНК, участвующие в
формировании митохондриальных рибосом, а так-же фенилаланиновая и валиновая
тРНК. Из длинных транскриптов вырезаются остальные тРНК и образуются
транслируемые мРНК, к 3'-концам которых при-соединяются полиадениловые
последовательности. 5'-концы этих мРНК не кэ-пируются, что необычно для
эвкариот. Сплайсинга (сращивания) не происхо-дит, поскольку ни один из
митохондриальных генов млекопитающих не содер-жит интронов.
|
ND1—ND6, ND4L — гены субъединиц НAД-H-дегидрогеназного комплекса; СОI—COIII — гены субъединиц цитохром-с-оксидазы; ATP6, ATP8 — гены субъединиц AТФ-синтетазы Cyt b — ген цитохрома b. |
Рис 4. Транскрипция мтДНК человека, содержащей 37 генов. Все транскрипты начинают синтезироваться в районе ori H. Рибосомные РНК вырезаются из длинного и короткого транскриптов Н-цепи. тРНК и мРНК образуются в результате процессинга из транскриптов обеих цепей ДНК. Гены тРНК обозначены светло-зеленым цветом.
Хотите узнать какие еще сюрпризы способен преподнести митохон-дриальный геном? Отлично! Читаем дальше!..
Несмотря на то, что в геномах митохондрий млекопитающих и дрожжей содержится приблизительно одинаковое количество генов, размеры дрожжево-го генома в 4-5 раз больше — около 80 тыс. пар нуклеотидов. Хотя кодирую-щие последовательности мтДНК дрожжей высоко гомологичны соответствую-щим последовательностям у человека, дрожжевые мРНК дополнительно имеют 5'-лидерную и 3'-некодирующую области, как и большинство ядерных мРНК. Ряд генов содержит еще и интроны. Так, в гене box, кодирующем цитохром-оксидазу b, имеется два интрона. Из первичного РНК-транскрипта автокатали-тически (без участия каких-либо белков) вырезается копия большей части пер-вого интрона. Оставшаяся РНК служит матрицей для образования фермента ма-туразы, участвующей в сплайсинге. Часть ее аминокислотной последовательно-сти закодирована в оставшихся копиях интронов. Матураза вырезает их, разру-шая свою собственную мРНК, копии экзонов сшиваются, и образуется мРНК для цитохромоксидазы b (рис.5). Открытие такого феномена заставило пере-смотреть представление об интронах, как о “ничего не кодирующих последова-тельностях”.
Рис 5. Процессинг (созревание) мРНК цитохромоксидазы b в митохондриях дрожжей. На первом этапе сплайсинга образуется мРНК, по которой синтезируется матураза, необходимая для второго этапа сплайсинга.
При изучении экспрессии митохон-дриальных генов Trypanosoma brucei
обнаружилось удивительное отклонение от одной из основных аксиом молекулярной
биологии, гласящей, что после-довательность нуклеотидов в мРНК в точности
соответствует таковой в коди-рующих участках ДНК. Оказалось, мРНК одной из
субъединиц цитохром-с-оксидазы редактируется,
т.е. после транскрипции изменяется ее первичная структура — вставляется четыре
урацила. В результате образуется новая мРНК, служащая матрицей для синтеза
дополнительной субъединицы фермента, пос-ледовательность аминокислот в которой
не имеет ничего общего с последова- тельностью, кодируемой
нередактированной мРНК (см. таблицу).
тельностью, кодируемой
нередактированной мРНК (см. таблицу).
Происходит это за счет сдвига рамки считыва-ния на число нуклеотидов, не кратное размеру триплета (в данном случае на четыре). Новая белковая субъединица, необходимая для работы фермента, об-разуется в митохондриях паразита только тогда, когда он попадает в организм холоднокровной мухи и нуждается в окислительном фосфорилировании для получения большого количества молекул АТФ. Если трипаносома живет в ор-ганизме теплокровных млекопитающих, ей достаточно АТФ, образующейся в процессе гликолиза. Впервые обнаруженное в митохондриях трипаносомы ре-дактирование РНК широко распространено в хлоропластах и митохондриях вы-сших растений. Найдено оно и в соматических клетках млекопитающих, напри-мер, в кишечном эпителии человека редактируется мРНК гена аполипопротеина.
Наибольший сюрприз ученым митохондрии преподнесли в 1979 г. До то-го времени считалось, что генетический код универсален и одни и те же трип-леты кодируют одинаковые аминокислоты у бактерий, вирусов, грибов, расте-ний и животных. Английский исследователь Беррел сопоставил структуру од-ного из митохондриальных генов теленка с последовательностью аминокислот в кодируемой этим геном субъединице цитохромоксидазы. Оказалось, что гене-тический код митохондрий крупного рогатого скота (как и человека) не просто отличается от универсального, он “идеален”, т.е. подчиняется следующему пра-вилу: “если два кодона имеют два одинаковых нуклеотида, а третьи нуклеоти-ды принадлежат к одному классу (пуриновых — А, Г, или пиримидиновых — У, Ц), то они кодируют одну и ту же аминокислоту”. В универсальном коде есть два исключения из этого правила: триплет АУА кодирует изолейцин, а кодон АУГ — метионин, в то время как в идеальном коде митохондрий оба эти трип-лета кодируют метионин; триплет УГГ кодирует лишь триптофан, а триплет УГА — стоп-кодон. В универсальном коде оба отклонения касаются прин-ципиальных моментов синтеза белка: кодон АУГ — инициирующий, а стоп-кодон УГА останавливает синтез полипептида. Идеальный код присущ не всем описанным митохондриям, но ни у одной из них нет универсального кода. Мож-но сказать, что митохондрии говорят на разных языках, но никогда — на языке ядра.
Различия между “универсальным” генетическим кодом и двумя митохондриальными кодами
|
Кодон |
Митохондриальный код млекопитающих |
Митохондриальный код дрожжей |
“Универсальный” код |
|
UGA |
Trp |
Trp |
Stop |
|
AUA |
Met |
Met |
Ile |
|
CUA |
Leu |
Thr |
Leu |
|
AGA AGG |
Cmon |
Arg |
Arg |
Как уже говорилось, в митохондриальном геноме позвоночных есть 22 ге-на тРНК. Каким же образом такой неполный набор обслуживает все 60 кодонов для аминокислот (в идеальном коде из 64 триплетов четыре стоп-кодона, в уни-версальном — три)? Дело в том, что при синтезе белка в митохондриях упроще-ны кодон-антикодонные взаимодействия — для узнавания используется два из трех нуклеотидов антикодона. Таким образом, одна тРНК узнает все четыре представителя кодонового семейства, отличающиеся только третьим нуклеоти-дом. Например, лейциновая тРНК с антикодоном ГАУ встает на рибосоме на-против кодонов ЦУУ, ЦУЦ, ЦУА и ЦУГ, обеспечивая безошибочное включе-ние лейцина в полипептидную цепь. Два других лейциновых кодона УУА и УУГ узнаются тРНК с антикодоном ААУ. В целом, восемь разных молекул тРНК узнают восемь семейств по четыре кодона в каждом, и 14 тРНК узнают разные пары кодонов, каждая из которых шифрует одну аминокислоту.
Важно, что ферменты аминоацил-тРНК-синтетазы, ответственные за при-соединение аминокислот к соответствующим тРНК митохондрий, кодируются в ядре клетки и синтезируются на рибосомах эндоплазматической сети. Таким образом, у позвоночных животных все белковые компоненты митохондриаль-ного синтеза полипептидов зашифрованы в ядре. При этом синтез белков в ми-тохондриях не подавляется циклогексимидом, блокирующим работу эвкариоти-ческих рибосом, но чувствителен к антибиотикам эритромицину и хлорамфени-колу, ингибирующим белковый синтез в бактериях. Этот факт служит одним из аргументов в пользу происхождения митохондрий из аэробных бактерий при симбиотическом образовании эвкариотических клеток.
Значение наличия собственной генетической системы для митохондрий
Почему митохондриям необходима собственная генетическая система, тогда как другие органеллы, например пероксисомы и лизосомы ее не имеют? Этот вопрос совсем не тривиален, так как поддержание отдельной генетической сис-темы дорого обходится клетке, если учесть необходимое количество дополни-тельных генов в ядерном геноме. Здесь должны быть закодированы рибосом-ные белки, аминоацил-тРНК-синтетазы, ДНК- и РНК-полимеразы, ферменты процессинга и модификации РНК и т. д. Большинство изученных белков из митохондрий отличаются по аминокислотной последовательности от своих аналогов из других частей клетки, и есть основание полагать, что в этих органе-ллах очень мало таких белков, которые могли бы встретиться еще где-нибудь. Это означает, что только для поддержания генетической системы митохондрий в ядерном геноме должно быть несколько десятков дополнительных генов.При-чины такого “расточительства” неясны, и надежда на то, что разгадка будет найдена в нуклеотидной последовательности митохондриальной ДНК, не опра-вдалась. Трудно представить себе, почему образующиеся в митохондриях бел-ки должны непременно синтезироваться именно там, а не в цитозоле.
