

 Реферат: Общее содержание воды в листьях калины в условиях биостанции
Реферат: Общее содержание воды в листьях калины в условиях биостанции
Парообразная влага. Представлена в почве в форме водяного пара и передвигается по градиенту абсолютной упругости пара; может также пассивно передвигаться с током воздуха. Содержание в почве парообразной влаги зависит от ряда факторов, в первую очередь от влажности, скважности и температуры почвенной среды. Эта форма почвенной влаги имеет некоторое значение в водоснабжении растений, т. к. парообразная влага при определенных температуре и давлении может конденсироваться и дополнять содержание в почве свободной или пленочной воды.
 |
Твердая влага. Эта форма воды присутствует в почве при снижении температуры до нуля градусов и ниже. Она неподвижна и недоступна для растений, но является резервом доступной влаги, которая возникает после таяния льда (Тарчевский, Жолкевич, 1989).
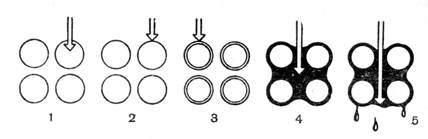
Рис. 2. Различные формы воды в почве: 1 – химически связанная вода; 2 – гигроскопически связанная вода; 3 – пленочная вода; 4 – капиллярная вода; 5 – гравитационная свободная вода; светлые кружки – частицы почвы (по Ф. Д. Сказкину).
1. 4. Распределение воды в растении
Содержание воды у разных растений различно. В листьях салата – 95%, кукурузы – 77 %. Количество воды неодинаково в разных органах растений. Приведем данные для подсолнечника: в листьях – 81 %, в стеблях – 88 %, в корнях – 71 %.
Содержание воды в листьях уменьшается обычно от морфологической верхней части к основанию стебля (ствола).
В вакуолях более старых клеток (нижний ярус) содержание воды больше, чем в молодых (верхний ярус). Большее количество воды в вакуолях клеток старых листьев объясняется увеличением их размеров с возрастом растения. Большая часть воды в клетках содержится в цитоплазме, а не в вакуоли (Сулейманов, 1974).
А) Вода в клеточных оболочках.
Содержание воды в клеточных стенках зависит от строения и ее химического состава. В оболочке зрелой клетки различают три слоя: срединную пластинку, первичную и вторичную оболочки. Первая из них состоит из пектата кальция, в составе второго слоя содержатся целлюлозные фибриллы, пропитанные пектиновыми веществами. Вторичная оболочка содержит целлюлозу, пектиновые вещества, лигнин, кутин, и она отлагается поверх первичной. Молекулы целлюлозы обладают гидрофильными свойствами, что связано, что связано с наличием гидроксильных групп. Всасывающая и водоудерживающая способность клеточной оболочки связана с пектиновыми веществами и ее целлюлозными компонентами. Лигнин может адсорбировать воду в количестве до 25 % от собственного веса. Гемицеллюлоза способна впитать воду до 100 % от сухого веса. В тургесцентных клетках большая часть воды удерживается, по-видимому, в свободных межфибриллярных пространствах. В указанных частях (клеточная стенка) тургосцентных клеток гидратной воды составляет менее 10 % от общего количества воды, содержащегося в клеточных оболочках. Клеточные стенки живых клеток способны удерживать значительно большие количества воды, чем ее компоненты, выделенные в чистом виде. В клетках с рыхлыми оболочками, с большими межфибриллярными пространствами количество воды больше, нежели в плотных клеточных стенках. Отложение инкрустирующих веществ приводит к уменьшению содержания воды в клеточной оболочке.
На содержание воды в клеточных стенках влияют различные вещества, находящиеся в ее соке и обладающие осмотическими свойствами. Имеет значение также концентрация сока, содержащегося в межфибриллярных пространствах.
Б) Воды, содержащаяся в проводящей системе растений
Проводящая система растений состоит из ксилемы и флоэмы (здесь имеют значение клетки-спутницы и клетки, выполняющие механические функции).
Ксилему, состоящую из сосудов, трахеид, волокон, древесной паренхимы и др. , можно рассматривать как распределительную систему, благодаря деятельности которой достигается снабжение различных органов растений водой.
Ткани, состоящие преимущественно из тонкостенных паренхимных клеток, например верхушечные и боковые меристемы, содержат очень много воды, часто свыше 90 %. В противоположность им толстостенные клетки или клетки, содержащие обильные отложения запасных веществ, в частности крахмала, отличаются в общем низким содержанием влаги. Молодые листья, содержащие относительно много цитоплазмы по сравнению с веществом клеточных оболочек, отличаются высоким содержанием воды (Козловский, 1969).
Особенно много воды в тканях, обладающих высокой физиологической активностью, наоборот, ее мало в тканях, имеющих небольшую физиологическую активность. Разумеется, указанный вопрос можно истолковать и по-другому, а именно, в тканях с малым содержанием воды физиологические процессы протекают менее интенсивно (Сулейманов, 1974).
1. 5. Содержание воды в растении
Вода, содержащаяся в растении, имеет двоякое происхождение: экзогенное и эндогенное. Большая часть ее, поступающая из почвы в растение через корневую систему, имеет экзогенное происхождение. Вода может быть поглощена растением в виде пара из атмосферного воздуха, но этот путь поступления не имеет существенного значения в обеспечении растений водой, так как это не может спасти их от гибели в условиях почвенной засухи.
Основным источником воды для растений является почва. Источники воды в почве – атмосферные осадки, грунтовая вода, поливная вода.
По отношению к воде растения подразделяются на три экологические группы: гидатофиты (водные), пойкилогидровые и гомеогидровые (Антипов, 1973). Указанные группы отличаются друг от друга по типу водообмена.
Гидатофиты. К этой группе относятся такие растения, у которых жизненные процессы протекают нормально лишь при непосредственном контакте их организма с внешней водой. Они не выдерживают сухости воздуха, быстро погибают при отсутствии непосредственного соприкосновения с водой. Вследствие того, что гидатофиты живут в воде, в их онтогенезе водообмен не изменяется. К гидатофитам относятся многие водоросли, некоторые грибы и отдельные лишайники, представители мохообразных, папоротникообразных и цветковых.
Из некоторых древних гидатофитов, перешедших на сушу, в результате адаптации к условиям недостатка воды, возникли пойкилогидровые низшие растения: наземные водоросли, многие грибы, лишайники.
У последних сохранились черты примитивной организации предков и примитивный водообмен. Об этом свидетельствует то, что они, подобно гидатофитам, поглощают воду всей своей поверхностью, не имеют специальных приспособлений для регулирования водообмена. Пойкилогидровые растения приспособились переносить обезвоживание, высыхание на них не действует губительно (Генкель, 1968). Свойствами пойкилогидровых растений обладают также и споры, семена высших растений. Они, также как и пойкилогидровые растения, не погибают при полном обезвоживании и не регулируют содержания воды. Они очень засухоустойчивые.
В онтогенезе пойкилогидровых растений водообмен сильно изменяется вследствие изменения содержания воды во внешней среде. Пойкилогидровые растения быстро поглощают воду и очень быстро ее испаряют. При полном насыщении указанные растения содержат мало связанной воды (Антипов, 1973). Пойкилогидровые растения приспособились к эффективному использованию воды, об этом свидетельствует резкое повышение интенсивности дыхания их после кратковременного намачивания.
К гомеогидровым растениям относится большинство папоротникообразных, цветковых и все голосеменные. При обезвоживании они погибают и они менее засухоустойчивы, нежели пойкилогидровые. Характерной особенностью гомеогидровых растений является то, что они имеют специальные ткани, органы для регулирования водообмена.
Сухопутные высшие растения также делятся на три экологические группы: гигрофиты, ксерофиты, мезофиты.
К гигрофитам относятся растения влажных местообитаний, они приспособлены к воздуху, насыщенному парами воды. К ним относятся: папоротники, фиалки, чистотел, болотный подмаренник и другие. У теневых гигрофитов устьица всегда открыты, интенсивность транспирации почти равна испарению свободной поверхности воды. Наблюдается выделение воды гидатодами. Световые гигрофиты распространены в жарком климате на почвах перенасыщенных водой и даже покрытых водой. Сюда относятся рис, культивируемый на залитых водой полях, болотные пальмы. Гигрофиты не выносят значительного водного дефицита, не обладают способностью приспосабливаться к почвенной засухе.
Ксерофиты – растения сухих местообитаний, они способны выносить продолжительную сухость воздуха и почвы, произрастают в областях с жарким и сухим климатом. Они встречаются на южном берегу Крыма, по берегам Средиземного моря и др. . Сюда относятся суккуленты – молочаи, алоэ, агавы, кактусы и др. . «Ксерофитами называются, - пишет П. А. Генкель (1946), - растения сухих местообитаний, обладающие способностью в процессе своего развития благодаря своим анатомо-физиологическим особенностям хорошо приспосабливаться к неблагоприятному влиянию атмосферной и почвенной засухи». Общим признаком почти всех ксерофитов является уменьшение их поверхности по сравнению с другими экологическими группами растений, что связывают с недостаточным снабжением их водой. Ксерофиты имеют небольшую узкую пластинку листа, у них листья мало или они не развиты, у некоторых видов листья сбрасываются в критический период, т. е. в условиях продолжительного недостатка воды. У многих растений листья свернуты в трубку или края их загнуты внутрь. Некоторые ксерофиты имеют многослойный эпидермис, сильно развитую кутикулу, последняя иногда покрыта восковым налетом. Ксерофиты отличаются многоклеточностью, плотным сложением, малым развитием межклетников, сильным развитием механических тканей.
Страницы: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17
