

 Статья: Генетическая рекомбинация в свете эволюции
Статья: Генетическая рекомбинация в свете эволюции
Статья: Генетическая рекомбинация в свете эволюции
Павел Михайлович Бородин, д.б.н., зав.лаб. рекомбинационного и сегрегационного анализа Ин-та цитологии и генетики СО РАН, проф. каф. цитологии и генетики НГУ.
Ничто в биологии не имеет смысла, кроме как в свете эволюции.
Федосий Добжанский
Рекомбинация - это процесс, который обеспечивает перемешивание генов в ряду поколений. При формировании половых клеток гены, полученные от родителей, “перетасовываются”, и в каждую гамету попадает только половина родительских генов. При оплодотворении гены двух родителей случайно комбинируются в зиготе. Сочетание этих двух случайных процессов - тасовки генов в генеративных клетках и встречи гамет - обеспечивает уникальность набора генов каждого организма.
Этот процесс был открыт в начале XX в. на основе анализа результатов скрещиваний. Сейчас в изучении рекомбинации используют весь арсенал современных методов молекулярной и клеточной биологии. И тем не менее процесс остается во многом загадочным. До сих пор идут бурные дебаты о том, зачем нужна рекомбинация. Непонятно, отчего она так сложно и, казалось бы, нелогично организована. Неясно, как распределяются по геному ее горячие и холодные точки. Попытаемся ответить на эти вопросы, рассмотрев рекомбинацию в свете эволюции.
Зачем нужна рекомбинация
Рекомбинация - главный генератор фенотипического разнообразия, того самого, с которым оперирует естественный отбор, тех отличий между организмами, которые играют решающую роль в их борьбе за существование. Мы привыкли думать, что эти различия определяются мутациями генов. Это и верно, и неверно одновременно.
Мутации меняют гены. Ген может быть неузнаваемо испорчен мутацией, изменен с сохранением функции (синонимически) или с ее потерей. Мы должны ясно понимать, что функция каждого гена определяется его взаимодействием с другими генами. Поэтому и функцию гена, и ее изменения следует рассматривать исключительно в рамках конкретного метаболического пути или регуляторной генной сети, в которых задействованы продукты этого гена. Бессмысленный или неверный ген из одной генной сети может приобрести новый, неожиданный смысл в другой; синоним в одном контексте оказаться антонимом в другом. Таким образом, мутации меняют фенотип не сами по себе, а в сочетании с другими генами.
Разнообразие фенотипов, которое мы наблюдаем, есть воплощенное разнообразие генных сочетаний. А поскольку рекомбинация обеспечивает постоянную генерацию все новых и новых сочетаний, мы имеем полное право назвать этот замечательный механизм генератором фенотипического разнообразия.
Рекомбинация, видимо, возникла одновременно или вскоре после появления жизни. Однако на первых порах она была робкой и спорадической. Такой она и остается в мире прокариот. Бактерии иногда входят в контакт друг с другом и обмениваются генетической информацией, чаще когда их жизнь становится хуже. Но из этого не следует, что рекомбинация непременно облегчает им жизнь, повышает их приспособленность. Она дает им шанс, надежду на то, что новая комбинация генов окажется полезной.
Регулярная, запланированная и обязательная рекомбинация появилась гораздо позже, одновременно или вскоре после возникновения эукариотических клеток. В пользу этого предположения свидетельствует тот факт, что у подавляющего большинства современных эукариот рекомбинация происходит регулярно, а ее молекулярные и клеточные механизмы у самых разных организмов поразительно сходны. Сходство мы обнаруживаем и в том, что у всех них рекомбинация так или иначе связана с размножением. У эукариот, в отличие от бактерий, результаты рекомбинации проявляются не у самих организмов, а у их потомков.
Если мы сравним размножение бесполых (не рекомбинирующих) и половых (регулярно рекомбинирующих) организмов, нам сразу бросится в глаза поразительная неэффективность последнего варианта размножения. Представим себе два острова. На одном живут самец и самка, способные к половому размножению и, следовательно, к рекомбинации. На другом - две самки, размножающиеся бесполым путем. Ограничим плодовитость и тех и других самок двумя потомками. После первого же цикла размножения на бесполом острове родится четыре потомка, а на половом - два. Если на половом острове оба родившихся детеныша будут одного пола, то на этом вся история закончится. Если на свет появятся самка и самец, то эта пара произведет еще двух потомков, а на бесполом острове их родится уже восемь. Таким образом, при заданных условиях численность популяции бесполого острова будет расти экспоненциально, а на половом она так и останется равной двум особям. Очевидно, что эффективность бесполого размножения значительно выше (рис.1).

Рис.1. Сравнение эффективности полового и бесполого размножения.
Почему же тогда у эукариот, как правило, размножение половое, а бесполое - лишь редкое исключение? Именно потому, что при половом размножении возможна рекомбинация. Но если организмы, размножающиеся половым путем, так значительно проигрывают бесполым в эффективности размножения, то рекомбинация должна давать им преимущества, с лихвой покрывающие этот гигантский проигрыш. В чем же они заключаются?
Вернемся на наши умозрительные острова. И на одном, и на другом острове в генеративных клетках их обитателей возникают мутации. Полностью защититься от мутаций в принципе невозможно, ведь с ними неизбежно сопряжено копирование ДНК. Большинство мутаций оказываются вредными. Парадоксально, но очень вредные мутации не так опасны для генофонда популяции, как не очень вредные. Очень вредные мутации несовместимы с жизнью, их носители немедленно выбраковываются, и, следовательно, такие мутации не накапливаются в генофонде. А не очень вредные передаются потомкам, затем у них возникают новые не очень вредные мутации, и в итоге генофонд бесполой популяции медленно, но верно деградирует (рис.2,а).
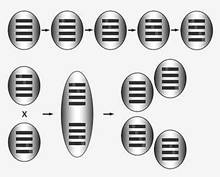
Рис.2. Накопление вредных мутаций (красные точки) при бесполом размножении (а) и их сортировка и элиминация при половом размножении (могут поддерживать свое существование) (б).
Выдающийся генетик Герман Мёллер впервые обратил внимание на медленную, но неуклонную деградацию бесполого генофонда за счет последовательного накопления не очень вредных мутаций. Сейчас в научной литературе этот процесс называется храповиком Мёллера. Мёллер показал, что бесполые популяции, несмотря на давление мутационного процесса, могут поддерживать свое существование за счет очень высокой численности и сильного давления стабилизирующего отбора, благодаря которому носители даже не очень вредных мутаций быстро погибают, а их место занимают клоны, свободные от мутаций.
Однако у храповика Мёллера есть еще одна неприятная особенность. Чем больше у организма генов, тем больше он накапливает мутаций. Вероятность мутации одного гена приблизительно равна 10-5 на гамету за поколение. Это значит, что каждая вторая из 10 тыс. гамет, содержащих 5 тыс. генов (именно столько их у бактерий), несет одну новую мутацию. Если в гамете 30 тыс. генов, как у нас млекопитающих, то каждая из 10 тыс. гамет несет в среднем три новых мутаций. Отсюда третье условие, позволяющее виду жить с храповиком Мёллера, - малый размер генома и как следствие - относительная простота организации.
Мощное и радикальное средство борьбы с храповиком Мёллера - рекомбинация. Перетасовывая гены при образовании гамет, она может перегрузить мутациями одни гаметы и одновременно недогрузить другие. В итоге особи, возникшие из перегруженных мутациями гамет, погибают, а продукты гамет, очищенных от мутаций, процветают (рис.2,б). Это позволяет рекомбинирующим организмам избавиться от ограничений, накладываемых храповиком Мёллера. Они могут позволить себе роскошь иметь большие геномы. Отсюда получается, что все мы высшие и сложные оттого, что наши далекие одноклеточные предки открыли для себя рекомбинацию и создали механизмы, гарантирующие регулярную перетасовку генов из поколения в поколение.
Гипотеза Мёллера - не единственное объяснение преимуществ рекомбинации. Очень подробные обзоры гипотез о преимуществах рекомбинации даны в книгах Дж.Мэнард Смита [1] и М.Ридли [2].
Как происходит рекомбинация
У всех эукариот рекомбинация происходит в мейозе [3]. Этот процесс изучают в школе, и по степени вызываемой у учащихся ненависти мейоз может соперничать разве что с двойным оплодотворением у растений по Навашину. Что обидно, поскольку процесс этот по-своему красив (рис.3).
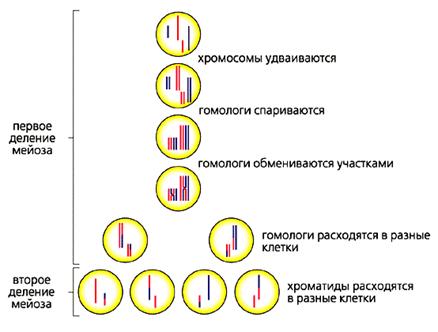
Рис.3. Схема мейоза.
Красным обозначены материнские хромосомы, синим - отцовские.
Некоторые сравнивают мейоз с танго [4]. В первом туре мейоза пары удвоенных гомологичных хромосом (одна получена от отца, другая от матери) находят друг друга, тесно свиваются, обмениваются участками, а потом расходятся в разные стороны, в разные клетки, чтобы уже не встретиться никогда. Второй тур мейоза менее романтичен. В нем гомологи выстраиваются вдоль экватора клетки и их хроматиды расходятся к разным полюсам. Так из одной генеративной клетки получаются четыре гаметы с разными наборами генов.
Рекомбинация происходит в первом, романтическом туре мейоза. Рассмотрим его более подробно. Сближение гомологичных хромосом начинается с того, что их концы, скользя по ядерной мембране, собираются в одной точке и формируется структура, романтически названная букетом. В нем гомологичные хромосомы оказываются поблизости друг от друга и приступают к взаимному опознанию. Оно, видимо, идет в два этапа: сначала приблизительное, а затем точное. Приблизительное опознание может происходить по принципу штрих-кодов. Известно, что хромосомы представляют собой комплекс ДНК с белками, набор которых и характер связывания с ДНК во многом определяются последовательностью нуклеотидов. Поэтому каждая хромосома отличается индивидуальным, только для нее специфичным распределением белков - штрих-кодом. Поскольку гомологичные хромосомы в основном сходны по последовательностям ДНК, они должны иметь сходные штрих-коды. Таким образом, грубое распознавание может быть достигнуто простым их совмещением.
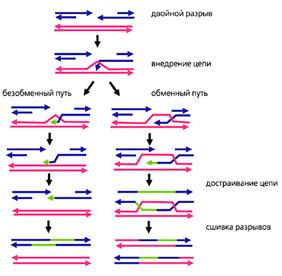
Рис.4. Молекулярные механизмы тонкого опознавания и рекомбинации гомологичных хромосом.
Красным обозначена ДНК материнской хромосомы, синим - отцовской, зеленым - ДНК, достроенная в ходе репарации разрывов.
Обратите внимание, что достройка идет по матрице ДНК гомолога, а не по собственной матрице [5].
Процесс тонкого опознания начинается с того, что в ДНК мейотических хромосом возникают множественные двунитевые разрывы (рис.4). Так, у мыши на этой стадии в каждой мейотической клетке образуется около 300 разрывов, а у лилии - несколько тысяч. В соматических клетках такое количество разрывов может появиться только в результате массированного облучения или обработки мощным мутагеном. Мейотическая клетка сама себе наносит эти повреждения с помощью белка Spo11 (гомолога топоизомеразы II архей). У архей нет мейоза, и этот белок участвует в репарации повреждений. Он разрезает поврежденную ДНК и воссоединяет свободные концы. У эукариот в мейозе белок Spo11 только режет, оставляя другим белкам заниматься воссоединением.
В воссоединении разорванных нитей ДНК активно участвует белок Rad51 (рис.5). У прокариот и в соматических клетках эукариот он задействован в репарации повреждений ДНК: в комплексе с другими белками связывается со свободными концами разорванных ДНК и внедряет их в ДНК гомологичных хромосом, одновременно расплетая ДНК-мишень. Задача внедренных участков состоит в том, чтобы найти комплементарные фрагменты определенной протяженности. К этому моменту гомологи уже прошли грубое выравнивание по штрих-коду, поэтому поиск происходит на относительно небольших расстояниях и именно в тех районах, где гомология наиболее вероятна. Найдя комплементарный участок, внедрившаяся нить ДНК спаривается с ним.

Рис.5. Мейотическая клетка обыкновенной бурозубки на стадии тонкого распознавания гомологов, окрашенная с использованием DAPI (специфичного для ДНК красителя), антител к рекомбинационному белку Rad51 и белку синаптонемного комплекса (SCP3) - осевого элемента мейотических хромосом.
Тонкое опознание заканчивается, когда количество связок между ДНК пары гомологичных хромосом достигает критического уровня. В мейотической клетке начинается разрезание связок. Большая часть разрезается и сшивается таким образом, что восстанавливается исходное состояние цепей ДНК (безобменный путь). Только небольшая их часть (у млекопитающих - меньше 0.1) сшивается крест-накрест (обменный путь), при этом ДНК одного из гомологов в пункте обмена соединяется с ДНК другого. Это и есть точки рекомбинации. Именно в них происходит переключение с одного гомолога на другой. В мейоз хромосомы вступили в таком виде, как они были получены от родителей, а выйдут из мейоза уже рекомбинантными. В материнской хромосоме будет часть отцовской, а в отцовской - часть материнской. Но это будут уже результаты рекомбинации, а мы пока вернемся к самому процессу.
Устранение опознавательных связок связано с нанесением новых повреждений в ДНК и репарацией этих повреждений. Эти процессы регулируются комплексом специфичных белков. Примечательно, что все они принадлежат к тем семействам белков, которые у прокариот и в соматических клетках эукариот участвуют в залечивании мутационных повреждений ДНК.
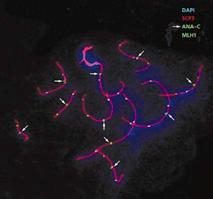
Рис.6. Мейотическая клетка обыкновенной бурозубки на стадии завершения рекомбинации, окрашенная с использованием DAPI, антител к белку репарации МLH1, центромерным белкам (ANA-C) и белку синаптонемного комплекса (SCP3).
Опознавательным знаком для точек рекомбинации служит белок MLH1 (рис.6). Он принадлежит к семейству белков мисматч репарации, функция которых - исправлять ошибки спаривания ДНК, т.е. устранять неспаренные нуклеотиды. Нас этот белок интересует в первую очередь как маркер точек рекомбинации. С помощью антител к MLH1, меченных флуоресцентными красителями, эти точки можно увидеть на хромосомах, а затем проанализировать частоту и распределение рекомбинационных событий по геному. Что мы и сделали на обыкновенной бурозубке. Для изучения мейоза она хороша тем, что у нее всего 10 пар хромосом, заметно отличающихся друг от друга по размеру и характеру дифференциального окрашивания [6]. Это позволило нам детально проанализировать особенности распределения точек рекомбинации по индивидуальным хромосомам.
Где происходит рекомбинация
Мы уже говорили, что рекомбинация - это случайный процесс перетасовки генов. Однако сами точки рекомбинации распределены по геному далеко не случайно (рис.7), а согласно ряду строгих правил.
Правило обязательного обмена гласит: всякая пара хромосом да имеет хотя бы один обмен. Общее число обменов на хромосому зависит от ее размера. На больших хромосомах бурозубки (af и bс) их может быть пять или шесть, но даже самые маленькие хромосомы (например, tu), как правило, имеют хотя бы один обмен.
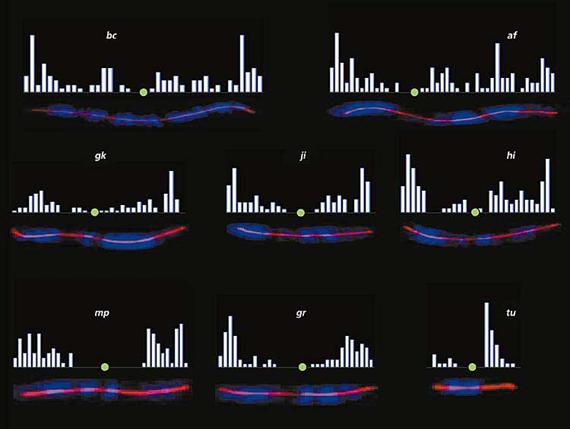
Рис.7. Распределение точек рекомбинации по хромосомам обыкновенной бурозубки.
Это правило обусловлено тем, что рекомбинация в мейозе, кроме генетического смысла (перетасовки генов), несет и чисто механическую функцию: образование и сохранение физической связи между гомологами вплоть до их расхождения в первом делении мейоза. Если между парой гомологов не произошло ни одного обмена, то они расходятся не как положено (один к одному полюсу деления, другой - к другому), а как попало, т.е. оба гомолога могут уйти в одному полюсу. Тогда у одних гамет будет избыточная доза генов, а у других этих генов не будет вовсе. И то, и другое чаще всего ведет к гибели организмов.
Правило теломерного пика: обмены могут быть в любых районах хромосом, но чаще всего они локализуются на самом краю хромосом - вблизи теломер. Это обусловлено, видимо, механикой рекомбинации, а не ее генетическим смыслом. Я уже говорил, что сближение хромосом начинается с того, что их концы собираются в одной точке на ядерной мембране, т.е. самый первый контакт между гомологами происходит именно на краях хромосом, и эти самые края находятся в тесном контакте дольше, чем все остальные районы. Там же возникают первые опознавательные связки на уровне ДНК, и времени на образование и разрешение этих связок отводится больше, чем в других точках хромосом.
Я с удовольствием вспоминаю эксперимент, который мы с И.П.Горловым провели еще в начале 1990-х годов [7]. Нас интересовал вопрос: что определяет частоту рекомбинации в каждом районе хромосомы - его положение или генетическое содержание? Мы сравнили распределение обменов на первой хромосоме у нормальных мышей и у гомозигот по инверсии терминальной половины этой хромосомы. Если важно положение, то распределение должно быть одинаковым; если важно содержание, то у гомозигот по инверсии и распределение должно быть инвертированным. Полученные нами распределения были одинаковыми (рис.8). Следовательно, решающую роль играет положение.

Рис.8. Распределение точек рекомбинации в хромосоме 1: слева - у нормальных мышей; в середине - у мышей, гомозиготных по инверсии в этой хромосоме, справа - распределение, которое было бы, если бы оно контролировалось не позицией, а генетическим содержанием района хромосомы.
Это механическое правило может иметь очень глубокий генетический и эволюционный смысл. Инверсии - одни из самых частых хромосомных перестроек, которые встречаются в природных популяциях и/или отличают виды друг от друга. Можно думать, что естественный отбор способствует фиксации инверсий, переносящих на края хромосом (т.е. в рекомбинационно горячие зоны) именно те гены, которые нужно часто перетасовывать, и наоборот, забрасывают в холодные зоны те блоки генов, которые лучше не трогать.
Правило интерференции: два обмена редко возникают в непосредственной близости друг от друга. Создается впечатление, что уже возникший обмен мешает (interfere) другому обмену возникнуть рядом. Это отталкивание обменов друг от друга получило название интерференции. Существует очень большой разброс в дистанциях между соседними обменами: от 1 мкм (т.е. на пределе разрешающей способности микроскопа) до 10 и более, а средние дистанции между соседними обменами у бурозубки составляют 5-6 мкм. Именно на этом расстоянии от теломерного пика обменов возникает второй пик. Если хромосома достаточно длинная, то примерно на таком же расстоянии возникнет третий пик, и так далее. Замечательно, что если мы обнаруживаем на большой хромосоме один-единственный обмен, то он, как правило, находится достаточно далеко от края. Почему? Потому что интерференция этого обмена не оставляет места для других обменов.
Парадоксально, но лучше всего явление интерференции интерпретируется в грубо механических терминах. Если представить хромосомы в виде упругих шлангов, скрученных друг с другом, становится понятно, что два перегиба рядом не возникают. Но в наше молекулярное время такие сантехнические аналогии выглядят по меньшей мере неприлично. Должны быть какие-то молекулярные механизмы интерференции обменов. По молекулярным понятиям несколько микрометров - это гигантское расстояние.
Существует несколько гипотез о механизмах интерференции, но ни одна из них не является общепринятой и ни одна из них мне не нравится. Поэтому я лучше приведу предложенную И.П.Горловым [8] гипотезу о том, зачем нужна интерференция. Суть ее такова. Ничего в природе не бывает бесплатно. Конечно, рекомбинация дает определенные преимущества, но она обходится и организму недешево. Чего стоит одно разрезание живой ДНК! Естественный отбор должен балансировать рекомбинацию таким образом, чтобы ее преимущества превышали затраты. Два обмена стоят дороже, чем один, где бы они ни были расположены. Но если мы подумаем о преимуществах, то станет понятно, что чем ближе они друг к другу, тем менее полезны в смысле перетасовки генов. Второй обмен в непосредственной близости от первого практически аннулирует его эффект, возвращая гомологичные хромосомы в исходное состояние (рис.9). Поэтому, если уж тратиться на обмены, то надо расположить их подальше друг от друга. Что и происходит благодаря интерференции, только до сих пор непонятно как.
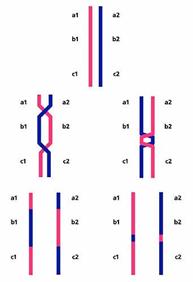
Рис.9. Результаты двух рекомбинационных событий на далеком (слева) и близком (справа) расстоянии друг от друга. В первом случае рекомбинантные хромосомы отличаются от родительских по набору генов a, b, c; во втором - не отличаются.
Все перечисленные выше правила так или иначе обусловлены позиционными эффектами, которые играют, видимо, главную роль в распределении обменов по хромосомам.
Правило светлого района связано с особенностями упаковки ДНК мейотических хромосом. Отдельные районы хромосом сильно отличаются по этому признаку. Хромосомы млекопитающих после обработки красителями, специфично связывающимися с ДНК, выглядят поперечно-полосатыми. В интенсивно окрашенных (темных) участках концентрация ДНК выше, чем в светлых. Особенности упаковки зависят от генетического состава ДНК. Сильно огрубляя ситуацию, можно сказать, что гены сосредоточены в основном в светлых районах, а темные главным образом состоят из бессмысленных и сильно повторенных последовательностей ДНК. Но это очень грубая картина. Гены есть и в темных районах, а в светлых довольно много бессмысленных последовательностей.
Правило светлого района состоит в том, что рекомбинация происходит в таких районах чаще, чем в темных. Однако пока не ясно почему. Можно думать, что ДНК светлых районов более активно участвует в поиске гомологии, в ней чаще возникают опознавательные связки и рекомбинационные обмены. На это указывает тот факт, что в мейотических хромосомах относительный размер светлых районов непропорционально больше, чем в митотических, а темных - непропорционально меньше. До наших экспериментов этот факт базировался на косвенных данных*. Было известно, что хромосомы, содержащие много светлых районов, в мейозе оказываются относительно более длинными, чем в митозе. Нам впервые удалось это увидеть, применив дифференциальное окрашивание мейотических хромосом обыкновенной бурозубки. Светлые районы были длиннее.
Правило светлого района на первый взгляд кажется нецелесообразным. Цель рекомбинации состоит в перетасовке генов. Но из этого не следует, что рекомбинация должна происходить именно в тех районах, где сконцентрированы гены. Гораздо логичней было бы производить обмены там, где генов нет. Результат был бы тем же, а риск повреждения генов за счет их разрезания, спаривания свободных цепей, подчистки результатов неверного спаривания был бы сведен к нулю. Однако логика эволюции не всегда совпадает с логикой инженера. Она (эволюция) никогда не создает механизмы с чистого листа. Она предпочитает слегка модифицировать уже существующие.
Мы уже говорили, что большинство белков, обеспечивающих рекомбинацию, - это гомологи бактериальных и эукариотических белков, участвующих в репарации, залечивании мутационных повреждений ДНК. Поскольку рекомбинация возникла в эволюции позже репарации и использует слегка измененную машину репарации, можно заключить, что она и произошла от репарации. Более того, скорее всего на первых этапах, когда еще не было мейоза, она была всего лишь одним из вариантов репарации, предназначенным для залечивания самых опасных и тяжелых повреждений ДНК - двунитевых разрывов. Если разорвана одна нить, разрыв можно залечить, используя вторую нить в качестве матрицы. Если же разорваны две нити, нужно найти гомологичный участок ДНК в другом месте генома, расплести его и использовать как матрицу (рис.4).
С этой точки зрения становится понятной и нецелесообразная на первый взгляд концентрация обменов в светлых, богатых генами районах хромосом, и кажущаяся избыточность разрывов ДНК на ранних стадиях точного опознавания гомологов, и тот факт, что только малая часть этих разрывов превращается в обмены, а большая - ни к каким обменам не ведет. В ходе подчистки необменных связок может происходить исправление потенциальных повреждений в генах.
Обратите внимание, однако, что репарация устраняет физические дефекты ДНК - разрывы, нарушения спаривания, полностью игнорируя смысл генетических текстов. Если повреждение обнаружено, например, в гене зеленых глаз, а в гомологичной хромосоме на этом месте находится ген карих глаз, то в восстановленной последовательности мы обнаружим именно его - ген карих глаз. Так благодаря рекомбинации один ген превращается в другой.
Рекомбинация - один из самых важных и самых загадочных генетических процессов. На первый взгляд он кажется слишком дорогим и ненужным, он происходит не так как надо и не там где надо. Но это только на первый взгляд. Этот процесс, как и все в биологии, приобретает смысл, если мы рассматриваем его в свете эволюции, пытаемся понять, как и из чего он возник, как менялся шаг за шагом, сохраняя старые функции и находя все новые применения.
В работе участвовали Н.М.Белоногова, Т.В.Карамышева, А.В.Поляков, М.И.Родионова и Н.Б.Рубцов, которым я искренне признателен.
Работа выполнена при поддержке Российского фонда фундаментальных исследований (проекты 01-04-48875 и 04-04-48024), ИНТАС и Программ Президиума РАН “Происхождение и эволюция жизни на Земле” и “Биоразнообразие и динамика генофондов”.
Список литературы
1. Мэйнард Смит Дж. Эволюция полового размножения. М., 1981.
2. Ridley M. The Red Queen: Sex and the Evolution of Human Nature. Penguin, 1995.
3. Богданов Ю.Ф. // Генетика. 2003. Т.39. С.453-473.
4. Roeder G.S. // Genes and Development. 1997. V.11. P.2600-2621.
5. Bishop D.K., Zickler D. // Cell. 2004. V.117. P.9-15.
6. Belonogova N.M., Karamysheva T.V., Biltueva L.S. et al. // Chromosome Res. 2006. V.14. P.673-679.
7. Gorlov I.P., Ladygina T.Yu., Serov O.L., Borodin P.M. // Heredity. 1991. V.66. P.453-458.
8. Gorlov I.P., Gorlova O.Y. // J. Theor. Biol. 2001. V.213. P.1-8.
