

 Шпаргалка: Меристемы или образовательные ткани
Шпаргалка: Меристемы или образовательные ткани
Клетки апикальных меристем более или менее изодиаметричны по размерам и многогранны по форме. Межклетников между ними нет, оболочки тонкие, содержат мало целлюлозы. Полость клетки заполнена густой цитоплазмой с относительно крупным ядром, занимающим центральное положение. Вакуоли многочисленные, мелкие, но под световым микроскопом обычно не заметны. Эргастические вещества, как правило, отсутствуют. Пластид и митохондрий мало и они мелки.
Клетки боковых меристем различны по величине и форме. Они примерно соответствуют клеткам тех постоянных тканей, которые из них в дальнейшем возникают. Так, в камбии встречаются как паренхимные, так и прозенхимные инициали. Из паренхимных инициалей образуется паренхима проводящих тканей, а из прозенхимных — проводящие элементы.
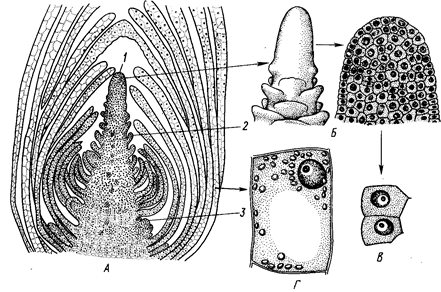
Рис. 1. Верхушечная меристема побега элодеи. А — продольный срез; 5 — конус нарастания (внешний вид и продольный срез); В — клетки первичной меристемы;
Г — паренхимная клетка сформировавшегося листа:
1 ~ конус нарастания, 2 — зачаток листа, 3 — бугорок пазушной почки
Зоны корня. Особенности морфологического строения каждой зоны в связи с выполняемыми функциями. Рост корня в длину и втолщину.
Наряду с побегом корень - основной орган высшего растения, в типичном случае выполняющий функцию минерального и водного питания. Другая важная функция корня, тесно связана с основной, - закрепление, растения в почве.
Различные части корня выполняют неодинаковые функции и характеризуются определенными морфологическими особенностями. Эти части получили название зон (рис. 2). Кончик корня снаружи всегда прикрыт корневым чехликом, защищающим апикальную меристему. Клетки корневого чехлика продуцируют слизь, покрывающую поверхность корня. Благодаря слизи снижается трение о почву, и ее частицы легко прилипают к корневым окончаниям и корневым волоскам.
Корневой чехлик состоит из живых паренхимных клеток, возникающих у большинства однодольных из особой меристемы, получившей название калиптрогена, а у двудольных и голосеменных — из верхушечной меристемы кончика корня. Водные растения корневого чехлика обычно не имеют.
Под чехликом располагается зона деления, представленная меристематической верхушкой корня, его апексом. В результате активности апикальной меристемы формируются все прочие зоны и ткани корня.
Делящиеся клетки сосредоточены в зоне деления, имеющей размеры около 1 мм. Эта часть корня заметно отличается от прочих зон своей желтоватой окраской. Вслед за зоной деления располагается зона растяжения (роста). Она также невелика по протяженности (несколько миллиметров), выделяется светлой окраской и как бы прозрачна. Клетки зоны роста практически не делятся, но способны растягиваться в продольном направлении, проталкивая корневое окончание в глубь почвы. Они характеризуются высоким тургором, что способствует активному раздвиганию частиц почвы. В пределах зоны роста происходит дифференциация первичных проводящих тканей.
Окончание зоны роста заметно по появлению на эпиблеме многочисленных корневых волосков. Корневые волоски располагаются в зоне—всасывания, функция которой понятна из ее названия. На корне она занимает участок от нескольких миллиметров до нескольких сантиметров. В отличие от зоны роста участки этой зоны уже не смещаются относительно частиц почвы. Основную массу воды и растворов солей молодые корни усваивают в зоне всасывания с помощью корневых волосков.
Корневые волоски появляются в виде небольших сосочков — выростов клеток эпиблемы. Рост волоска осуществляется у его верхушки. Оболочка корневого волоска растягивается быстро. По прошествии определенного времени корневой волосок отмирает. Продолжительность его жизни не превышает 10—20 дней.
Выше зоны всасывания, там, где исчезают корневые волоски, начинается зона проведения. Строение этой зоны на разных ее участках неодинаково. По этой части корня вода и растворы солей, поглощенные корневыми волосками, транспортируются в вышележащие отделы растения. Участки зоны проведения занимают фиксированное положение относительно участков почвы, не смещаясь относительно их. Несмотря на фиксированное положение в пространстве конкретных участков зон поглощения и проведения, эти зоны сдвигаются вследствие верхушечного роста. В результате этого всасывающий аппарат постоянно перемещается в почве.
В пределах одной корневой системы существуют корни, выполняющие разные функции. У большинства растений различаются ростовые и сосущие окончания. Ростовые окончания долговечнее, относительно мощные, быстро удлиняются и продвигаются в глубь почвы. Сосущие окончания недолговечны, возникают в большом числе на вростовых корнях
и удлиняются медленно. У деревьев и кустарников выделяют видоизмененные скелетные и полускелетные корни, на которых возникают недолговечные корневые мочки, несущие множество сосущих окончаний.
|
|
|
|
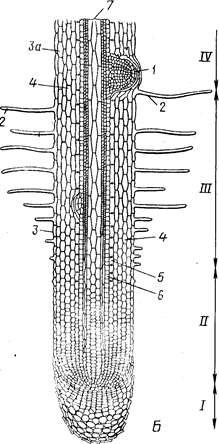
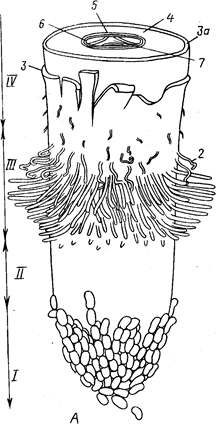
Рис.2 Общий вид {А) и продольный срез (5) корневого окончания (схема). 1— корневой чехлик; 11 - зона роста и растяжения; ///— зона корневых волосков, или зона всасывания; IV— начало зоны проведения (в этой зоне также закладываются боковые корни):
1- заложившийся боковой корень, 2 — корневые волоски на эпиблеме, 3 — эпиблема, За — экзодерма, 4 — первичная кора,
5 — эндодерма, 6 — перицикл, 7 — осевой цилиндр
Факторы среды и процессы роста и развития. Фитогормоны. Фотопериодизм. Факторы среды и процессы роста и развития.
Внешние факторы, или факторы среды, также оказывают заметный эффект на рост и развитие. Рассмотрим здесь главнейшие факторы — свет, тепло и влагу. Свет оказывает глубокое влияние на внешнее строение растений, причем его действие многообразно. Свет влияет на дыхание и прорастание семян, образование корневищ и клубней, формирование цветков, на листопад, переход почек в состояние покоя. Растения, выращенные при отсутствии света (этиолированные растения), обгоняют в росте растения, выросшие на свету. Интенсивное освещение нередко усиливает процессы дифференцировки.
Для каждого растения существует свой температурный оптимум роста и развития. Температурные минимумы для роста и развития в среднем лежат в интервале 5 —15°С, оптимумы — при 25 — 35°С, максимумы — в пределах 45 — 55°С. Низкие и высокие температуры могут нарушать покой семян, почек и сделать возможным их прорастание и распускание. Образование цветков — это переход из вегетативного состояния в генеративное. Индуцирование, т. е. ускорение этого процесса холодом, называют яровизацией. Без процесса яровизации многие растения (свекла, репа, сельдерей, озимые злаки) не способны к цветению.
Огромное значение для роста, прежде всего в фазе растяжения, имеет обеспеченность водой. Недостаток воды влечет за собой мелкоклеточность, а следовательно, отставание в росте.
Фитогормоны — это химические факторы, вырабатываемые в крайне малых количествах, но способные давать значительный физиологический эффект. Фитогормоны, выработанные в одной части растения, транспортируются в другую часть, вызывая там соответствующие изменения в зависимости от генной модели воспринимающей клетки.
Известны три класса фитогормонов, действующих по преимуществу как стимуляторы: ауксины, гиббереллины и цитокинины. Два класса гормонов — абсцизовая кислота и этилен — оказывают в основном тормозящее действие. Механизм действия различных гормонов неодинаков.
Многие процессы обмена веществ, роста, развития и движения подвержены ритмическим колебаниям. Иногда эти колебания следуют смене дня и ночи (циркадные ритмы), иногда связаны с длиной дня (фотопериодизм). Пример ритмических движений — ночное закрывание цветков, опускание и продольное складывание листьев, раскрытых и приподнятых в дневное время. Эти движения связаны с неравномерным тургором. Во многих случаях эти процессы контролируются внутренней хронометрической системой — физиологическими часами, по-видимому, существующими у всех эукариотических организмов. У растений важнейшая функция физиологических часов — регистрация длины дня и вместе с тем времени года, что определяет переход к цветению или подготовку к зимнему покою (фотопериодизм).
Виды, растущие на севере (севернее 60°с.ш.), должны быть преимущественно длиннодневными, поскольку их короткий вегетационный период совпадает с продолжительной длиной дня. В средних широтах (35—40° с. ш.) встречаются растения как длиннодневные, так и короткодневные. Здесь весенние или осеннецветущие виды относятся к короткодневным, а цветущие в разгар лета — к длиннодневным.
Фотопериод имеет большое значение для характера распространения растений. В процессе естественного отбора у видов генетически закрепилась информация о длине дня своих местообитаний и об оптимальных сроках начала цветения, Даже у растений, размножающихся вегетативно, длина дня определяет соотношение между сезонными изменениями и накоплением запасных веществ. Виды, индифферентные к длине дня, являются потенциальными осмополитами. Сюда же относятся и виды, цветущие с ранней весны до поздней осени. Другие виды не могут выходить за пределы географической широты, определяющей их способность к цветению при соответствующей длине дня. Фотопериодизм важен и в практическом отношении, поскольку он определяет возможности продвижения южных растений на север, а северных — на юг.
