

 Реферат: Структурная и молекулярная организация генного вещества
Реферат: Структурная и молекулярная организация генного вещества
Минорные РНК. По мере изучения компонентов живой клетки и отдельных фракций тотальной клеточной РНК выяснялось, что тремя главными видами РНК дело не ограничивается. Оказалось, что в природе существует множество других видов РНК. Это, в первую очередь, так называемые "малые РНК", которые содержат до 300 нуклеотидов, часто с неизвестными функциями. Как правило, они ассоциированы с одним или несколькими белками и представлены в клетке в виде рибонуклеопротеидов - "малых РНП" [8].
Малые РНК присутствуют во всех отделах клетки, включая цитоплазму, ядро, ядрышко, ми-тохондрии. Большая часть тех малых РНП, функции которых известны, участвует в механизмах посттранскрипционной обработки главных видов РНК (RNA processing) - превращении предшественников мРНК в зрелые мРНК (сплайсинг), редактировании мРНК, биогенезе тРНК, созревании рибосомных РНК. Один из наиболее богато представленных в клетках видов малых РНП (SRP) играет ключевую роль в транспорте синтезируемых белков через клеточную мембрану. Известны виды малых РНК, выполняющих регуляторные функции в трансляции. Специальная малая РНК входит в состав важнейшего фермента, ответственного за поддержание редупликации ДНК в поколениях клеток - теломеразы. Следует сказать, что их молекулярные размеры сопоставимы с размерами клеточных глобулярных белков. Таким образом, постепенно становится ясно, что функционирование живой клетки определяется не только многообразием синтезируемых в ней белков, но и присутствием богатого набора разнообразных РНК, из которых малые РНК в значительной мере имитируют компактность и размеры белков.
Рибозимы. Вся активная жизнь построена на обмене веществ - метаболизме, и все биохимические реакции метаболизма происходят с надлежащими для обеспечения жизни скоростями только благодаря высокоэффективным специфическим катализаторам, созданным эволюцией. На протяжении многих десятилетий биохимики были уверены, что биологический катализ всегда и всюду осуществляется белками, называемыми ферментами, или энзимами. И вот в 1982-1983 гг. было показано, что в природе имеются виды РНК, которые, подобно белкам, обладают высокоспецифической каталитической активностью [9, 10]. Такие РНК-катализаторы были названы рибозимами. Представлению об исключительности белков в катализе биохимических реакций пришел конец.
В настоящее время рибосому тоже принято рассматривать как рибозим. Действительно, все имеющиеся экспериментальные данные свидетельствуют о том, что синтез полипептидной цепи белка в рибосоме катализируется рибосомной РНК, а не рибосомными белками. Идентифицирован каталитический участок большой рибосомной РНК, ответственный за катализ реакции транспептидации, посредством которой осуществляется наращивание полипептидной цепи белка в процессе трансляции.
Вирусные РНК. Кроме нуклеиновых кислот (ДНК и РНК), организующих и обслуживающих жизнь клеточных организмов, в природе существуют паразитические молекулы ДНК и РНК. Одетые в защитную белковую оболочку, они называются вирусами. Соответственно, вирусы подразделяются на ДНК-содержащие и РНК-содержащие. Собственно в самих вирусных частицах никакой жизни нет - это просто способ упаковки, консервации и распространения внеклеточного генетического вещества. При попадании в живую клетку вирусная белковая оболочка сбрасывается, а генетическое вещество - нуклеиновая кислота - начинает функционировать как паразит, направляя жизнь клетки на синтез белков, ею кодируемых, и на репликацию самой себя. Так называемое "размножение" вирусов в клетке есть производство многочисленных копий вирусной ДНК или РНК путем репликации, с последующим их "одеванием" в оболочку из синтезированных клеткой вирусных белков.
Что касается репликации вирусных ДНК, то ее механизм мало чем отличается от редупликации генетического материала - ДНК - самой клетки. В случае же вирусных РНК реализуются процессы, которые подавлены или вовсе отсутствуют в нормальных клетках, где вся РНК синтезируется только на ДНК как на матрице. При инфекции РНК-содержащими вирусами ситуация может быть двоякой. В одних случаях на вирусной РНК как на матрице синтезируется ДНК ("обратная транскрипция"), а уж на этой ДНК транскрибируются многочисленные копии вирусной РНК. В других, наиболее интересных для нас случаях на вирусной РНК синтезируется комплементарная цепь РНК, которая и служит матрицей для синтеза - репликации - новых копий вирусной РНК. Таким образом при инфекции РНК-содержащими вирусами реализуется принципиальная способность РНК детерминировать воспроизведение своей собственной структуры, как это имеет место у ДНК.
К РНК-содержащим вирусам примыкает другая группа молекулярных паразитов - вироиды. Это патогенные РНК, не содержащие и не кодирующие никаких белков, но тоже способные к репликации в живых системах. Тем самым вирусные и вироидные РНК демонстрируют способность РНК не только кодировать белки, но и служить полноценным воспроизводящимся генетическим материалом. Вирусы и вироиды часто рассматриваются как эволюционные реликты, и процесс репликации РНК без участия ДНК может отражать очень ранний этап эволюции жизни, когда ДНК еще не утвердилась в качестве специализированной формы хранения и воспроизведения генетической информации в поколениях клеток.
Мультифункциональность РНК. Суммирование и обзор знаний о функциях РНК позволяют говорить о необыкновенной многофункциональности этого полимера в живой природе. Можно дать следующий список основных известных функций РНК.
• Генетическая репликативная функция: структурная возможность копирования (репликации) линейных последовательностей нуклеотидов через комплементарные последовательности. Функция реализуется при вирусных инфекциях и аналогична главной функции ДНК в жизнедеятельности клеточных организмов - редупликации генетического материала.
• Кодирующая функция: программирование белкового синтеза линейными последовательностями нуклеотидов. Это та же функция, что и у ДНК. И в ДНК, и в РНК одни и те же триплеты нуклеотидов кодируют 20 аминокислот белков, и последовательность триплетов в цепи нуклеиновой кислоты есть программа для последовательной расстановки 20 видов аминокислот в полипептидной цепи белка.
• Структурообразующая функция: формирование уникальных трехмерных структур. Компактно свернутые молекулы малых РНК принципиально подобны трехмерным структурам глобулярных белков, а более длинные молекулы РНК могут образовывать и более крупные биологические частицы или их ядра.
• Функция узнавания: высокоспецифические пространственные взаимодействия с другими макромолекулами (в том числе белками и другими РНК) и с малыми лигандами. Эта функция, пожалуй, главная у белков. Она основана на способности полимера сворачиваться уникальным образом и формировать специфические трехмерные структуры. Функция узнавания является базой специфического катализа.
• Каталитическая функция: специфический катализ химических реакций рибозимами. Данная функция аналогична энзиматической функции белков-ферментов.
В целом РНК предстает перед нами столь удивительным полимером, что, казалось бы, ни времени эволюции Вселенной, ни интеллекта Творца не должно было бы хватить на ее изобретение. Как можно было видеть, РНК способна выполнять функции обоих принципиально важных для жизни полимеров - ДНК и белков. Неудивительно, что перед наукой и встал вопрос: а не могло ли возникновение и самодостаточное существование мира РНК предшествовать появлению жизни в ее современной ДНК-белковой форме?
ПРОИСХОЖДЕНИЕ ЖИЗНИ
Белково-коацерватная теория Опарина. Пожалуй, первая научная, хорошо продуманная теория происхождения жизни абиогенным путем была предложена биохимиком А.И. Опариным еще в 20-х годах прошлого века [11, 12]. Теория базировалась на представлении, что все начиналось с белков, и на возможности в определенных условиях спонтанного химического синтеза мономеров белков - аминокислот - и белковоподобных полимеров (полипептидов) абиогенным путем. Публикация теории стимулировала многочисленные эксперименты в ряде лабораторий мира, показавшие реальность такого синтеза в искусственных условиях. Теория быстро стала общепринятой и необыкновенно популярной.
Основным ее постулатом было то, что спонтанно возникавшие в первичном "бульоне" белковоподобные соединения объединялись" в коацерватные капли - обособленные коллоидные системы (золи), плавающие в более разбавленном водном растворе. Это давало главную предпосылку возникновения организмов - обособление некой биохимической системы от окружающей среды, ее компартментализацию. Так как некоторые белковоподобные соединения коацерватных капель могли обладать каталитической активностью, то появлялась возможность прохождения биохимических реакций синтеза внутри капель - возникало подобие ассимиляции, а значит, роста коацервата с последующим его распадом на части - размножением. Ассимилирующий, растущий и размножающийся делением коацерват рассматривался как прообраз живой клетки (рис. 5).
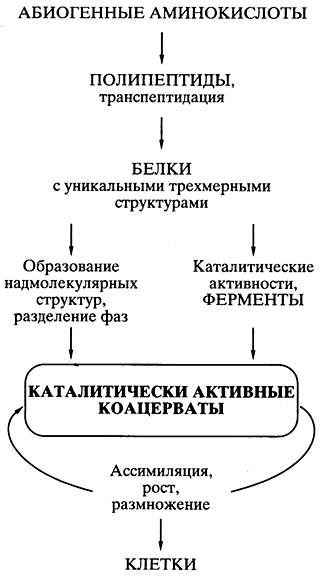
Рис. 5. Схематическое
представление пути происхождения жизни
согласно белково-коацерватной теории А.И. Опарина
Все было хорошо продумано и научно обосновано в теории, кроме одной проблемы, на которую долго закрывали глаза почти все специалисты в области происхождения жизни. Если спонтанно, путем случайных безматричных синтезов в коацервате возникали единичные удачные конструкции белковых молекул (например, эффективные катализаторы, обеспечивающие преимущество данному коацервату в росте и размножении), то как они могли копироваться для распространения внутри коацервата, а тем более для передачи коацерватам-потомкам? Теория оказалась неспособной предложить решение проблемы точного воспроизведения - внутри коацервата и в поколениях - единичных, случайно появившихся эффективных белковых структур.
