

 Реферат: Мир прокариотной клетки
Реферат: Мир прокариотной клетки
Рост и способы размножения
Под ростом прокариотной клетки понимают согласованное увеличение количества всех химических компонентов, из которых она построена. Рост является результатом множества скоординированных биосинтетических процессов, находящихся под строгим регуляторным контролем, и приводит к увеличению массы (а следовательно, и размеров) клетки. Но рост клетки не беспределен. После достижения определенных (критических) размеров клетка подвергается делению.
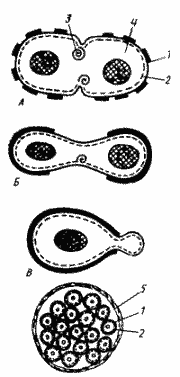
Для подавляющего большинства прокариот характерно равновеликое бинарное поперечное деление, приводящее к образованию двух одинаковых дочерних клеток. При таком способе деления имеет место симметрия в отношении продольной и поперечной оси. У большинства грамположительных эубактерий и нитчатых цианобактерий деление происходит путем синтеза поперечной перегородки, идущего от периферии к центру (рис. 20, А). Так у Bacillus subtilis в середине клетки сначала имеет место кольцевое впячивание ЦПМ, сопровождающееся формированием мезосом разного внешнего вида. Они образуются в месте закладки поперечной перегородки, и предполагается их активное участие в процессах синтеза пептидогликана и других компонентов клеточной стенки. Поперечная перегородка формируется из ЦПМ и пептидогликанового слоя, ее наружные слои синтезируются позднее. Клетки большинства грамотрицательных эубактерий делятся путем перетяжки. У Е. coli на месте деления обнаруживается постепенно увеличивающееся и направленное внутрь искривление ЦПМ и клеточной стенки (рис. 20, 5). Синтез новой клеточной стенки может происходить в нескольких местах или только в зоне формирования поперечной перегородки (рис. 20, А, Б).
Вариантом бинарного деления является почкование, которое можно рассматривать как неравновеликое бинарное деление. При почковании на одном из полюсов материнской клетки образуется маленький вырост (почка), увеличивающийся в процессе роста. Постепенно почка достигает размеров материнской клетки, после чего отделяется от последней. Клеточная стенка почки полностью синтезируется заново (рис. 20, В). В процессе почкования симметрия наблюдается в отношении только продольной оси. При равновеликом бинарном делении материнская клетка, делясь, дает начало двум дочерним клеткам и сама, таким образом, исчезает. При почковании материнская клетка дает начало дочерней клетке, и между ними можно в большинстве случаев обнаружить морфологические и физиологические различия: есть старая материнская клетка и новая дочерняя. В этом случае можно наблюдать процесс старения. Так, для некоторых штаммов Rhodomicrobium показано, что материнская клетка способна отпочковывать не более 4 дочерних клеток. Дочерние клетки лучше приспосабливаются к меняющимся условиям. Почкование обнаружено в разных группах прокариот: среди фото- и хемотрофов, осуществляющих авто- и гетеротрофный конструктивный метаболизм. Вероятно, оно в процессе эволюции возникало несколько раз.
|
Рис. 20. Способы деления и синтез клеточной стенки у прокариот: А — деление путем образования поперечной перегородки; Б — деление путем перетяжки; В — почкование; Г — множественное деление: 1 — клеточная стенка (толстой линией обозначена клеточная стенка материнской клетки, тонкой — заново синтезированная); 2 — ЦПМ; 3 — мембранная структура; 4 — цитоплазма, в центре которой расположен нуклеоид; 5 — дополнительный фибриллярный слой клеточной стенки |
Бинарное деление может происходить в одной или нескольких плоскостях. В первом случае, если после деления клетки не расходятся, это приводит к образованию цепочек палочковидных или сферических клеток, во втором — к клеточным скоплениям разной формы (см. рис. 3, 4–6). Расхождение образовавшихся дочерних клеток происходит в результате лизиса среднего слоя клеточной стенки.
Для одной группы одноклеточных цианобактерий описано размножение путем множественного деления. Оно начинается с предварительной репликации хромосомы и увеличения размеров вегетативной клетки, которая затем претерпевает ряд быстрых последовательных бинарных делений, происходящих внутри дополнительного фибриллярного слоя материнской клеточной стенки. Это приводит к образованию мелких клеток, получивших название баеоцитов12, число которых у разных видов колеблется от 4 до 1000. Освобождение баеоцитов происходит путем разрыва материнской клеточной стенки (рис. 20, Г). Таким образом, в основе множественного деления лежит принцип равновеликого бинарного деления. Отличие заключается в том, что в этом случае после бинарного деления не происходит роста образовавшихся дочерних клеток, а они снова подвергаются делению.
Деление прокариотной клетки начинается, как правило, спустя некоторое время после завершения цикла репликации молекулы ДНК. Вероятно, репликация бактериальной хромосомы запускает какие-то процессы, ведущие к клеточному делению. Более детальное изучение у разных видов прокариот взаимосвязи между репликацией ДНК и делением клетки не привело к однозначным результатам. Получены данные о том, что сигналом к клеточному делению служит начало репликации ДНК, ее завершение или репликация определенного локуса бактериальной хромосомы. Таким образом, в норме существует вполне определенная временная связь между репликацией хромосомы и делением бактериальной клетки. Воздействия различными химическими веществами и физическими факторами, приводящие к подавлению репликации ДНК, останавливают и клеточное деление. Однако при некоторых условиях связь между обоими процессами может быть нарушена, и клетки способны делиться в отсутствие синтеза ДНК. Это удалось получить введением определенных мутаций в генетический аппарат бактериальной клетки.
Нарушить последовательность процессов репликации бактериальной хромосомы и клеточного деления также можно, выращивая бактерии при разной температуре. Культивирование Bacillus subtilis на богатой питательной среде при 37° приводит к интенсивному делению бактериальной хромосомы и росту клеток, в результате чего в культуре образуются нитевидные клетки, содержащие множество хромосомных копий с отсутствующими совсем или недосформированными (незамкнутыми) поперечными перегородками. При замедлении скорости роста наблюдается деление нитевидных клеток, приводящее к образованию бактериальных клеток нормальной длины.
Внутрицитоплазматические включения
В цитоплазме прокариот обнаруживаются различные включения. Одни из них следует рассматривать как активно функционирующие структуры, другие — как продукты клеточного метаболизма, не выделяющиеся наружу, но откладывающиеся внутри клетки. Некоторые цитоплазматические включения имеют явно приспособительное значение. И наконец, многие из них являются запасными веществами, отложение которых клеткой происходит в условиях избытка питательных веществ в окружающей среде, а потребление наблюдается, когда организм попадает в условия голодания.
12 Баеоцит — по-гречески маленькая клетка.
К числу внутрицитоплазматических включений, выполняющих определенную функцию в фотосинтезе, относятся хлоросомы зеленых бактерий и фикобилисомы цианобактерий. В этих структурах локализованы пигменты, поглощающие кванты света и передающие их в реакционные центры, т. е. выполняющие роль антенны. Хлоросомы имеют форму продолговатых пузырьков длиной 90—150 и шириной 25—70 нм, окруженных однословной электронно-плотной мембраной толщиной 2—3 нм, построенной только из белка. Они располагаются в непосредственной близости от ЦПМ, плотно к ней примыкая (см. рис. 4). В хлоросомах локализованы бактериохлорофиллы с, d или е. Водорастворимые пигменты белковой природы (фикобили-протеины) цианобактерий содержатся в особых структурах — фикобилисомах, расположенных правильными рядами на внешних поверхностях фотосинтетических мембран и под электронным микроскопом имеющих вид гранул диаметром 28—55 нм (см. рис. 4).
В клетках некоторых прокариот из групп фототрофных и хемолитотрофных эубактерий содержатся структуры, имеющие форму многогранника с 4—6 сторонами и диаметром 90— 500 нм, получившие название карбоксисом, или полиэдральных тел (см. рис. 4). Под электронным микроскопом удалось показать, что они заполнены гранулярным содержимым и окружены однослойной мембраной белковой природы толщиной примерно 3 нм. Карбоксисомы состоят из частиц рибулозодифосфаткарбоксилазы, фермента, катализирующего фиксацию СО2 на рибулозодифосфате в восстановительном пентозофосфатном цикле. До настоящего времени окончательно не выяснено, в какой форме находится фермент в карбоксисомах: в инертном или функционирующем состоянии. Имеются данные в пользу того, что в активно растущей культуре больше фермента находится в растворимой форме. При переходе в стационарную фазу увеличивается доля рибулозодифосфаткарбоксилазы в составе карбоксисом. Эти данные указывают на возможную роль карбоксисом как структур, обеспечивающих защиту фермента от воздействия внутриклеточными протеазами и, таким образом, его консервирование.
Примером внутрицитоплазматических включений, имеющих приспособительное значение, служат магнитосомы и газовые вакуоли, или аэросомы, обнаруженные у водных прокариот. Газовые вакуоли найдены у представителей, относящихся к 15 таксономическим группам. Это сложно организованные структуры, напоминающие пчелиные соты (см. рис. 4). Состоят из множества регулярно расположенных газовых пузырьков, имеющих форму вытянутого цилиндра с заостренными концами (диаметр 65—115, длина 200—1200 нм). Каждый пузырек окружен однослойной белковой мембраной толщиной 2—3 нм, построенной из одного или двух видов белковых молекул, и заполнен газом, состав которого идентичен таковому окружающей среды. Мембрана газовых пузырьков проницаема для газов, но не проницаема для воды. Число газовых пузырьков, составляющих аэросому, у разных видов различно и зависит от внешних условий. Основная функция газовых вакуолей состоит в обеспечении плавучести водных организмов, которые с их помощью могут регулировать глубину, выбирая более благоприятные условия. При увеличении объема и числа газовых пузырьков плотность цитоплазмы уменьшается, и клетки перемещаются в верхние слои воды. Сжатие газовых пузырьков, наоборот, приводит к погружению клеток. За несколькими исключениями, газовые вакуоли присущи безжгутиковым видам. Их, вероятно, можно рассматривать как альтернативу жгутикам для движения в вертикальной плоскости.
Запасные вещества прокариот представлены полисахаридами, липидами, полипептидами, полифосфатами, отложениями серы (см. рис. 4; табл. 5). Из полисахаридов в клетках откладываются гликоген, крахмал и крахмалоподобное вещество— гранулеза. Последняя — специфический запасной полисахарид анаэробных споровых бактерий группы клостридиев. Названные полисахариды построены из остатков глюкозы. В неблагоприятных условиях они используются в качестве источника углерода и энергии.
Таблица 5. Запасные вещества прокариот
| Запасное вещество | Структурные характеристики | Химический состав | Функции | Распространение |
| Гранулы гликогена (a-гранулы) | сферической формы, диаметр 20–100 нм | высокомолекулярные полимеры глюкозы | источник углерода и энергии | широко распространенный тип запасных веществ |
| Гранулы поли-b-оксимасляной кислоты | диаметр 100–1000 нм; окружены однослойной белковой мембраной 2–3 нм толщиной | 98% полимера поли-b-оксимасляной кислоты, 2% белка | источник углерода и энергии | широко распространены только у прокариот |
| Цианофициновые гранулы | размер и форма различны; могут достигать в диаметре 500 нм |
полипептид, содержащий аргинин и аспарагиновую кислоту (1:1), мол. масса — 25–100x103 Да |
источник азота | обнаружены у многих видов цианобактерий |
| Гранулы полифосфата | диаметр приблизительно 500 нм, зависит от объекта и условий выращивания | линейные полимеры ортофосфата | источник фосфора и, возможно, энергии | распространенный тип запасных гранул |
| Гранулы серы | диаметр 100–800 нм; окружены однослойной белковой мембраной толщиной 2–3 нм | включения жидкой серы | донор электронов или источник энергии |
пурпурные серобактерии, бесцветные бактерии, окисляющие H2S |
| Углеводородные гранулы | диаметр 200–300 нм; окружены белковой оболочкой 2–4 нм толщиной | углеводороды того же типа, что и в среде | источник углерода и энергии |
представители родов Arthrobacter, Acinetobacter, Mycobacterium, Nocardia и другие прокариоты, использующие углеводороды |
Липиды накапливаются в виде гранул, резко преломляющих свет и поэтому хорошо различимых в световой микроскоп. Запасным веществом такого рода является полимер b-оксимасляной кислоты, накапливающийся в клетках многих прокариот. У некоторых бактерий, окисляющих углеводороды, поли-b-оксимасляная кислота составляет до 70% сухого вещества клеток. Отложение липидов в клетке происходит в условиях, когда среда богата источником углерода и бедна азотом. Липиды служат для клетки хорошим источником углерода и энергии.
Другой широко распространенный тип запасных веществ многих прокариот — полифосфаты, содержащиеся в гранулах, называемых волютиновыми, или метахроматиновыми, зернами. Используются клетками как источник фосфора. Полифосфаты содержат макроэргические связи и, таким образом, являются депо энергии, хотя считается, что их роль как источника энергии незначительна.
Специфическим запасным веществом цианобактерий являются цианофициновые гранулы. Химический анализ показал, что они состоят из полипептида, содержащего аргинин и аспарагиновую кислоту в эквимолярных количествах. Остов молекулы построен из остатков аспарагиновой кислоты, соединенных пептидными связями, а к ее b-карбоксильным группам присоединены остатки аргинина. Для синтеза цианофицина необходимы затравка, молекулы АТФ, ионы К+ и Mg2+. Процесс не закодирован в иРНК и не связан с рибосомами. Появление цианофициновых гранул при культивировании цианобактерий в среде с азотом и их исчезновение при истощении среды по азоту указывают на то, что они в клетке служат резервом азота, мобилизуемым при его недостатке в среде.
Для прокариот, метаболизм которых связан с соединениями серы, характерно отложение в клетках молекулярной серы. Сера накапливается, когда в среде содержится сероводород, и окисляется до сульфата, когда весь сероводород среды оказывается исчерпанным. Для аэробных тионовых бактерий, окисляющих H2S, сера служит источником энергии, а для анаэробных фотосинтезирующих серобактерий она является донором электронов.
Обращает внимание, что все запасные вещества представлены в виде высокомолекулярных полимерных молекул, в ряде случаев отграниченных от цитоплазмы белковой мембраной, т. е. находятся в осмотически неактивном состоянии. Это важно, так как в противном случае сосредоточение в цитоплазме большого числа молекул осмотически активных веществ оказало бы на клетку отрицательное действие.
ЛИТЕРАТУРА
- Влавианос-Арванитис А., Олескин А.В. Биополитика. Био-окружение. Био-силлабус. Афины: Биополитическая Интернациональная Организация. 1993.
- Горелов А.А. Социальная экология. М. Изд-во Ин-та философии РАН. 1998.
- Гусев М.В. К обсуждению вопроса об антропоцентризме и биоцентризме// Вест. Моск. ун-та. Сер. 16 (Биология). 1991. N 1. С.3--6.
- Данилова Н.Н., Крылова А.Л. Физиология высшей нервной деятельности. М.: Учебная литература. 1997.
- Дерягина М.А. Эволюционная антропология. М.: Изд-во УРАО. 1999.
- Дерягина М.А, Бутовская М.Л. Этология приматов. М.: МГУ. 1992.
- Дольник В.Р. Непослушное дитя биосферы. Беседы о человеке в компании птиц и зверей. М.: Педагогика. 1994.
- Дольник В.Р. Вышли мы все из природы. Беседы о поведении человека в компании птиц, зверей и детей. М.: Linka Press. 1996.
- Дьюсбери Д. Поведение животных. Сравнительные аспекты. М.: Мир. 1981.
- Захаров А.А. Организация сообществ у муравьев. М.: Наука. 1991.
- Зорина З.А., И.И. Полетаева, Ж.И. Резникова. Основы этологии и генетики поведения. М.: Изд-во МГУ. 1999.
- Карпинская Р.С., Лисеев И.К., Огурцов А.П.. Философия природы: коэволюционная стратегия. М.: Интерпракс. 1995. С. 13--78.
- Ламсден Ч., Гуршурст А. Генно-культурная коэволюция: человеческий род в становлении // Человек. 1991. № 3. С.11--22.
- Лоренц К.З. Агрессия (так называемое зло). М.: Прогресс. 1994.
- Майерс Д. Социальная психология. Спб., М., Харьков, Минск: Питер. 2000.
Мак-Фарленд Д. Поведение животных. Этология и психобиология. М.: Мир. 1988.
