

 Реферат: Менструальный цикл и его регуляция
Реферат: Менструальный цикл и его регуляция
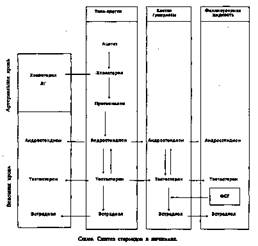
Этапы развития доминантного фолликула: Примордиальный фолликул состоит из яйцеклетки, окруженной одним рядом уплощенных клеток фолликулярного эпителия (рис. 14). В процессе созревания фолликула яйцеклетка увеличивается в размере, клетки фолликулярного эпителия размножаются и округляются, образуется зернистый слой фолликула (stratum granulosum). В гранулезных клетках зреющего фолликула имеются рецепторы
к гонадотропным гормонам, определяющие чувствительность яичников к гонадотропинам и регулирующие процессы фолликуло- и стероидогенеза. В толще зернистой оболочки за счет секреции и распада клеток фолликулярного эпителия и транссудата из кровеносных сосудов появляется жидкость. Яйцеклетка оттесняется жидкостью к периферии, окружается 17—50 рядами клеток гранулезы. Возникает яйценосный холмик (cumulus oophorus). В граафовом пузырьке яйцеклетка окружена стекловидной оболочкой (zona pellucida). Строма вокруг зреющего фолликула дифференцируется на наружную (tunica externa thecae folliculi) и внутреннюю покрышки фолликула (tunica interna thecae follicula). Зреющий фолликул превращается в зрелый.
В фолликулярной жидкости резко увеличивается содержание эстрадиола (Ез) и фолликулостимулирующего гормона. Подъем уровня Е2 стимулирует выброс лютеинизирующего гормона и овуляцию. Фермент коллагеназа обеспечивает изменения в стенке фолликула (истончение и разрыв). Играют роль в разрыве преовуляторного фолликула простагландины (ПГРао и гага) и протеолитические ферменты, содержащиеся в фолликулярной жидкости, а также окситоцин и релаксин.
На месте разорвавшегося фолликула образуется желтое тело, клетки которого секретируют прогестерон, эстрадиол и андрогены. Полноценное желтое тело образуется только тогда, когда в преовуляторном фолликуле содержится достаточное количество гранулезных клеток с высоким содержанием рецепторов ЛГ.
Стероидные гормоны продуцируются клетками гранулезы, клетками theca folliculi interna и, в меньшей степени, клетками theca folliculi externa. Клетки гранулезы и тека-клетки участвуют в синтезе эстрогенов и прогестерона, а клетки theca folliculi externa —в синтезе андрогенов (см. схему).
Исходным материалом для всех стероидных гормонов является холе-стерол, образующийся из ацетата или липопротеидов низкой плотности. Он поступает в яичник с током крови. В синтезе стероидов на первых этапах участвуют ФСГ и ЛГ, ферментные системы — ароматазы. Андрогены синтезируются в тека-клетках под влиянием ЛГ и с током крови попадают в гранулезные клетки. Конечные этапы синтеза (превращения андрогенов в эстрогены) происходит под влиянием ферментов.
В клетках гранулезы образуется белковый гормон — ингибин, тормозящий выделение ФСГ. В фолликулярной жидкости, желтом теле, матке и маточных трубах обнаружен окситоцин. Окситоцин, секретируемый яичником, оказывает лютеолитическое действие, способствуя регрессу желтого тела. Вне беременности в клетках гранулезы и желтого тела очень мало релаксина, а в желтом теле при беременности его содержание возрастает во много раз. Релаксин оказывает токолитическое действие на матку и способствует овуляции.
III уровень —передняя доля гипофиза (аденогипофиз). В аденоги-пофизе секретируются гонадотропные гормоны: фолликулостимулирующий, или фоллитропин (ФСГ), лютеинизирующий, или лютропин (ЛГ); пролак-тин (ПрЛ); другие тройные гормоны: тиреотропный гормон, тиротропин (ТТГ); соматотропный гормон (СТГ); адренокортикотропный гормон, кор-тикотропин (АКТГ); меланостимулирующий, меланотропин (МСГ) и липо-тропный (ЛПГ) гормоны. ЛГ и ФСГ являются гликопротеидами, ПрЛ — полипептидом.
 Железой-мишенью для ЛГ и ФСГ является
яичник. ФСГ стимулирует рост фолликула, пролиферацию клеток гранулезы,
образование рецепторов ЛГ на поверхности клеток гранулезы. ЛГ стимулирует
образование андрогенов в тека-клетках. ЛГ И ФСГ способствуют овуляции. ЛГ
стимулирует
Железой-мишенью для ЛГ и ФСГ является
яичник. ФСГ стимулирует рост фолликула, пролиферацию клеток гранулезы,
образование рецепторов ЛГ на поверхности клеток гранулезы. ЛГ стимулирует
образование андрогенов в тека-клетках. ЛГ И ФСГ способствуют овуляции. ЛГ
стимулирует
синтез прогестерона в лютеинизированных клетках гранулезы после овуляции.
Основная роль пролактина—стимуляция роста молочных желез и регуляция лактации. Он оказывает гипотензивное действие, дает жиромоби-лизующий эффект. Повышение уровня пролактина тормозит развитие фолликулов и стероидогенез в яичниках.
IV уровень репродуктивной системы — гипофизотропная зона гипоталамуса: вентромедиальные, дорсомедиальные и аркуатные ядра. В этих ядрах образуются гипофизотропные гормоны. Выделен, синтезирован и описан рилизинг-гормон — люлиберин. Выделить и синтезировать фоллибе-рин до настоящего времени не удалось. Поэтому гипоталамические гона-дотропные либерины обозначают ГТ-РГ, так как рилизинг-гормон стимулирует выделение как ЛГ, так и ФСГ передней долей гипофиза.
ГТ-РГ гипоталамуса из аркуатных ядер по аксонам нервных клеток попадает в терминальные окончания, тесно соприкасающиеся с капиллярами медиальной возвышенности гипоталамуса. Капилляры формируют портальную кровеносную систему, объединяющую гипоталамус и гипофиз. Особенностью этой системы является возможность тока крови в обе стороны, что
важно в осуществлении механизма обратной связи. Нейросекрет гипоталамуса оказывает биологическое действие на организм различными путями.
Основной путь — парагипофизарный — через вены, впадающие в синусы твердой мозговой оболочки, а оттуда в ток крови. Трансгипофизарный путь —через систему воротной вены к передней доле гипофиза. Обратное влияние на гипоталамус (стероидный контроль половых органов) осуществляется через вертебральные артерии. Секреция ГТ-РГ генетически запрограммирована и происходит в определенном пульсирующем ритме с частотой примерно один раз в час. Этот ритм получил название цирхорального (часового). Он формируется в пубертатном возрасте и является показателем зрелости нейросекреторных структур гипоталамуса. Цирхоральная секреция ГТ-РГ запускает гипотоламо-гипофизарно-яичниковую систему. Под влиянием ГТ-РГ происходит выделение ЛГ и ФСГ из передней доли гипофиза.
В модуляции пульсации ГТ-РГ играет роль эстрадиол. Величина выбросов ГТ-РГ в преовуляторный период (на фоне максимального выделения эстрадиола) значительно выше, чем в раннюю фолликулиновую и лютей-новую фазы. Частота выбросов остается прежней. В дофаминергических нейронах аркуатного ядра гипоталамуса есть рецепторы эстрадиола.
Основная роль в регуляции выделения пролактина принадлежит дофа-минергическим структурам гипоталамуса. Дофамин (ДА) тормозит выделение пролактина из гипофиза. Антагонисты дофамина усиливают выделение пролактина.
V уровень в регуляции менструального цикла — надгипоталамиче-ские церебральные структуры. Воспринимая импульсы из внешней среды и от интерорецепторов, они передают их через систему передатчиков нервных импульсов (нейротрансмиттеров) в нейросекреторные ядра гипоталамуса.
В эксперименте показано, что в регуляции функции гипоталамических нейронов, секретирующих ГТ-РГ, ведущая роль принадлежит дофамину, норадреналину и серотонину. Функцию нейротрансмиттеров выполняют ней-ропептиды морфиноподобного действия (опиоидные пептиды) — эндорфины (ЭНД) и энкефалины (ЭНК). Они регулируют гонадотропную функцию гипофиза. ЭНД подавляют секрецию ЛГ, а их антагонист — налоксон — приводит к резкому повышению секреции ГТ-РГ. Считают, что эффект опиоидов осуществляется за счет изменения содержания ДА (ЭНД снижают синтез ДА, вследствие чего стимулируется секреция и выделение пролактина).
В регуляции менструального цикла участвует кора большого мозга. Имеются данные об участии амигдалоидных ядер и лимбической системы в нейрогуморальной регуляции менструального цикла. Электрическое раздражение амигдалоидного ядра (в толще больших полушарий) вызывает в эксперименте овуляцию. При стрессовых ситуациях, при перемене климата, ритма работы наблюдаются нарушения овуляции. Нарушения менструального цикла реализуются через изменение синтеза и потребления нейротрансмиттеров в нейронах мозга.
Таким образом, репродуктивная система представляет собой суперсистему, функциональное состояние которой определяется обратной связью входящих в нее подсистем. Регуляция внутри этой системы может идти по длинной петле обратной связи (гормоны яичника —ядра гипоталамуса; гормоны яичника—гипофиз); по короткой петле—передняя доля гипофиза — гипоталамус; по ультракороткой —ГТ-РГ—нервные клетки гипоталамуса. Обратная связь может быть как отрицательной, так и положительной. При низком уровне эстрадиола в раннюю фолликулярную фазу усиливается выделение ЛГ передней долей гипофиза — отрицательная обратная связь.
Овуляторный пик выделения эстрадиола вызывает выброс ФСГ и ЛГ-положительная обратная связь. Примером ультракороткой отрицательной связи может служить увеличение секреции ГТ-РГ при снижении его концентрации в нейросекреторных нейронах гипоталамуса.
Помимо циклических изменений в системе гипоталамус — гипофиз— яичники и в органах-мишенях на протяжении менструального цикла имеют место циклические изменения функционального состояния многих систем («менструальная волна»). Эти циклические изменения у здоровых женщин находятся в пределах физиологических границ.
При изучении функционального состояния центральной нервной системы выявлена некоторая тенденция к преобладанию тормозных реакций, снижение силы двигательных реакций во время менструаций.
В фазе пролиферации отмечается преобладание тонуса парасимпатического, а в секреторной фазе — симпатического отделов вегетативной нервной системы. Состояние сердечно-сосудистой системы в течение менструального цикла характеризуется волнообразными функциональными колебаниями. Так, в I фазе менструального цикла капилляры несколько сужены, тонус всех сосудов повышен, ток крови быстрый. Во II фазе менструального цикла капилляры несколько расширены, тонус сосудов снижен; ток крови не всегда равномерный.
