Реферат: Генная инженерия
Реферат: Генная инженерия
Реферат: Генная инженерия
СОДЕРЖАНИЕ
Введение. 2
ГЛАВА 1. Теоретические предпосылки формирования генной инженерии как науки. 3
1.1. Открытие двойной структуры ДНК и матричного синтеза. 3
1.2.РЕСТРИКТАЦИОННЫЕ ЭНДОНУКЛЕАЗЫ. 5
1.3.ПРИНЦИПЫ ТЕХНОЛОГИЙ РЕКОМБИНАНТНЫХ ДНК. 5
1.4. ИДЕНТИФИКАЦИЯ И АНАЛИЗ ГЕНОВ. 8
1.5. ГИБРИДИЗАЦИЯ НУКЛЕИНОВЫХ КИСЛОТ. 9
1.6. СОРТИРОВКА ХРОМОСОМ. 10
1.7. СЕКВЕНИРОВАНИЕ ДНК. 11
1.8.ДИНАМИЧНОСТЬ ГЕНОМА. 12
ГЛАВА 2. ВОЗМОЖНОСТИ ГЕННОЙ ИНЖЕНЕРИИ. 14
2.1. ЧТО БУДЕТ СДЕЛАННО ПОСЛЕ ЗАВЕРШЕНИЯ АНАЛИЗА ГЕНОМА ЧЕЛОВЕКА. 18
Глава 3. Области практического применения генной инженерии. 19
3.1. Создание трансгенных растений. 19
3.1.1 Изменение свойств сельскохозяйственных технических растений 21
3.1.2. Генетическая модификация пластид. 22
3.3. ГЕННЫЕ ВАКЦИНЫ 22
3.2.1. Актуальность разработки новых вакцин 22
3.2.2.Разработка ДНК-вакцин 24
3.2.3. Повышение эффективности и безопасности иммунизации 28
3.2.4. Упрощение разработки и производства новых вакцин 30
3.2.5. Упрощение требований к условиям хранения 31
3.2.6. Вопросы безопасности применения 33
3.2.7. Участие фармацевтических компаний в разработке ДНК-вакцин 33
3.3. Генотерапия 35
3.3.1. Методы генетической трансфекции в генной терапии. 38
Глава 4. Перспективы клонирования животных 40
Введение.
Генная инженерия - направление исследований в молекулярной биологии и генетике, конечной целью которых является получение с помощью лабораторных приемов организмов с новыми, в том числе и не встречающимися в природе, комбинациями наследственных свойств. В основе генной инженерии лежит обусловленная последними достижениями молекулярной биологии и генетики возможность целенаправленного манипулирования с фрагментами нуклеиновых кислот. К этим достижениям следует отнести установление универсальности генетического кода, то есть факта, что у всех живых организмов включение одних и тех же аминокислот в белковую молекулу кодируются одними и теми же последовательностями нуклеотидов в цепи ДНК ; успехи генетической энзимологии, предоставившей в распоряжение исследователя набор ферментов, позволяющих получить в изолированном виде отдельные гены или фрагменты нуклеиновой кислоты, осуществлять in vitro синтез фрагментов нуклеиновых кислот, объединить в единое целое полученные фрагменты. Таким образом, изменение наследственных свойств организма с помощью генной инженерии сводится к конструированию из различных фрагментов нового генетического материала, введение этого материала в рецепиентный организм, создания условий для его функционирования и стабильного наследования.
ГЛАВА 1. Теоретические предпосылки формирования генной инженерии как науки.
1.1. Открытие двойной структуры ДНК и матричного синтеза.
Начальные работы американских учёных Уотсона и Крика были произведены в 1953 году. Они дали возможность развиваться генной инженерии в качестве самостоятельного раздела науки. Эти открытия заключены в следующем:
Была открыта двойная структура ДНК и постулирован её матричный синтез. Двойная спираль ДНК при репликации разделится и вдоль нити ДНК, специальные ферменты-полимеры, собирают точные копии материнской ДНК, таким образом в клетке перед делением две совершенно одинаковые молекулы ДНК, одна из которых после деления клетки попадает в дочернюю клетку. Таким образом дочерняя клетка несет ту же самую информацию, что и материнская, следовательно выполняет те же самые функции. Итак, в клетках живого организма возможен особый тип реакции – матричный синтез. Одна молекула – матрица, а вторая строится по её программе. репликация ДНК синтез всех видов РНК и сборка молекул белка, в соответствии со структурой и-РНК – это все варианты матричного синтеза, который происходит всегда при участии нуклеиновых кислот.
По тому же самому механизму осуществляется сборка РНК, только не двух спиралей, а одной. Этот процесс получил название – транскрипция. Поток информации в клетке обеспечивает реакции матричного синтеза: репликация ДНК(необходима для передачи наследственной информации дочерним клеткам), транскрипция(синтез и-РНК в ядре клетки) и трансляция(сборка белковой цепи на и-РНК при помощи рибосомы).
Казалось бы, что на рубеже 70-х годов молекулярная биология достигла определённой степени завершенности: были установлены структура и механизм репликации ДНК, провозглашена «центральная догма» экспрессии гена (транскрипция и трансляция), выявлены основные аспекты регуляции активности гена. В этот период главным объектом молекулярно-генетических исследований были микроорганизмы. Переход к эукариотам(включая человека) встретился с дополнительными проблемами и трудностями, и кроме того, существовавшие в то время методы не позволяли рассчитывать на получение принципиально новых результатов. Стремительный порыв в развитии молекулярной генетики в начале 70-х годов стал благодаря появлению нового экспериментального инструмента – рестриктационных эндонуклеаз. Был открыт путь для широкомасштабного получения генных продуктов(физически значимых белков) и для генетического манипулирования с различными организмами. Наши знания о структуре генетического материала и эукариот, в разных областях таких: как действие гена, популяционная генетика, эволюция и генетическая консультация, включая пренатальную диагностику. Достигнутые успехи заставили ученых задуматься об этической стороне манипулирования с человеческим зародышем, об возникновения возбудителей различных болезней в процессе генно-инженерных исследований. Многие из этих вопросов были подняты самими учеными активно работающих в данной области. В настоящее время большинство исследователей считали, что опасения касающиеся, генной инженерии, не имеют достаточно оснований, но многие этические проблемы остаются нерешенными и продолжают возникать новые.
В прошлом генетика и медицинская генетика развивалась как относительно независимые отрасли науки, теперь многие из их разделов оказались вовлечённые в общее русло молекулярно-генетических исследований, и провести между ними грань – трудно.
Сейчас, множество ученых заняты различными работами связанные с проблемами генной инженерии – это и методы, основанные на использовании рестриктационных ферментов, анализ гена человека, методы гибридизации нуклеиновых кислот, секвенирование ДНК, сортировки хромосом при помощи цитофиурометрииии и многое, многое другое. Попытаюсь дать необходимые разъяснения по важнейшим работам из этого ряда.а
Начнём с условий, которым должен соответствовать ген человека, что бы получить полную характеристику его структуры:
1) соответствующие фрагменты ДНК должны быть идентифицированы однозначно.
2) они должны быть выделены и накоплены в количестве, должностном для биохимического анализа.
3) должна быть определена вся нуклеотидная последовательность.
Принципы, на которых основаны эти три метода, кратко будут описаны ниже. Мы начнем с описания второго, поскольку прогресс в выделении и клонировании генов был решающим для развития новой генетики.
1.2.РЕСТРИКТАЦИОННЫЕ ЭНДОНУКЛЕАЗЫ.
_ґ_ьї0’чїяяяяёПb_\Jчї_____
различные штаммы E-coli, Арбер обнаружил, что ДНК этого фага при переходе через бактерию разрезается и теряет свою инфекционность. Оказалось, что ни классические рекомбинационные процессы, ни мутации в этом не участвуют. Более того, такая судьба постигала не только фаговую, но и любую чужеродную ДНК, попадающую в бактерию. Такое разрезание(рестрикцию) следует рассматривать как защитный механизм клетки. Как было показано в дальнейшем, рестриктация чужеродной ДНК осуществляется ферментами, называемыми рестриктационными эндонуклеазами(рестриктазами). Встаёт вопрос, почему рестриктазы не разрезают ДНК собственной клетки? Ответ был найден Арбером и состоял в следующем: эти ферменты вступают в реакцию с определенными участками в ДНК, так называемыми сайтами узнавания, которые в клетке защищены метильными группами(метилированы). Правда, первые из открытых эндонуклеаз не были специфическими, а действовали случайным образом. Первой рестриктазой, которая расщепляла ДНК, в стого определенном месте, была Hind, открытая Смитом в конце 60-х годов. Этот фермент впервые использован Натсоном и соавторами для создания рестриктационнй карты генома вируса SO40. Берг уловил особое свойство двухцепочной ДНК формировать при обработке рестриктазами так называемые «липкие концы».После разрезания одна из цепей оказывается длиннее, чем другая, на несколько нуклеотидов.Эти нуклеотиды могут свободно спариваться с другими, например с комплиментарными нуклеотидами другого фрагмента ДНК с «липкими концами». Благодаря этому, ДНК из различных источников может объединяться, образуя рекомбинантные молекулы.
1.3.ПРИНЦИПЫ ТЕХНОЛОГИЙ РЕКОМБИНАНТНЫХ ДНК.
Было выделено много рестриктаз(более 150),расщепляющих ДНК в специфических сайтах. Например эндонуклеаза R1 регистрирует двухцепочную ДНК по двум сайтам таким образом, что образуются два липких конца:
G-A-A-T-T-C
||| || || || || |||
C-T-T-A-A-G
Липкие концы различных молекул ДНК, расщеплённых этим ферментом, могут вступать по четырём –A-T-парам. Рестриктационные эндонуклеазы различаются по тем сайтам ДНК, которые они распознают и разрезают. Их можно использовать для различных целей. Однако наиболее распространенным этапом является их применение для амплификации специфической определения нуклеотидных последовательностей фрагментов ДНК, необходимо для ДНК или для изучения механизмов экспрессии генов. Последняя проблема наиболее важна в практическом аспекте: гены контролирующие образование функционально активных белков, теперьможно вводить в бактерии и размножать(амплифицировать).эта процедура называется клонированием генов. Благодаря ей, появилась возможность вырабатывать в больших количествах белки, которые раньше удавалось получить ничтожно мало. Эта технология основана на следующем принцепе: помимо своей собственной кольцевой хромосомы, бактерии часто содержат дополнительные маленькие кольцевидные молекулы двух цепочной ДНК,называемые плазмидами.
Плазмиды реплицируются автономо и сами могут содержать гены, определяющие устойчивость бактерий к антибиотикам или контролирующие синтез веществ, например: колицинов, убивающих другие бактерии(см. рис.1).
Плазмидную ДНК можно выделить и ращепить подходящей рестриктазой только в одном сайте, превратив кольцевую молекулу в линейную с липкими концами.
Фрагменты любой чужеродной ДНК с такими же липкими концами(полученными
после разрезания аналогичной рестриктазой) можно сшить с плазмидой ДНК с помощью лигазы.
Р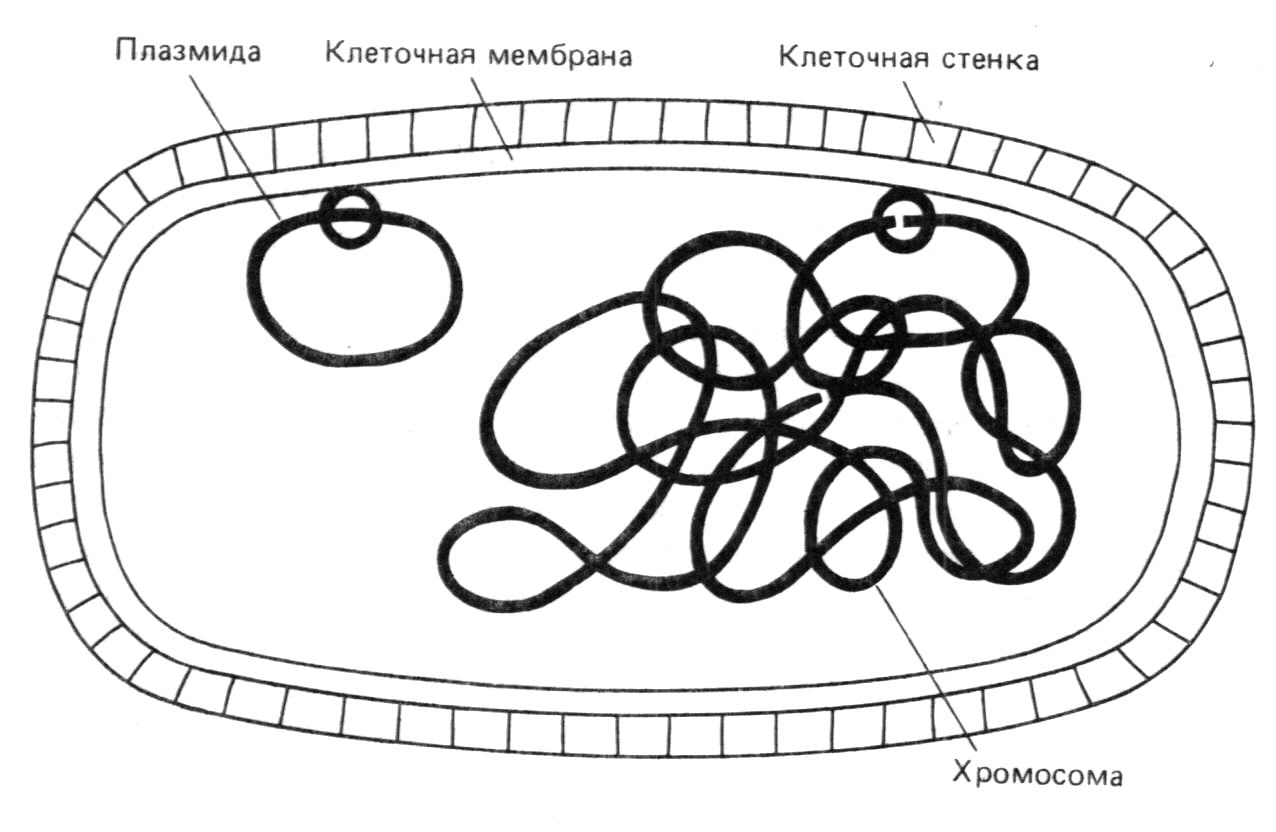
ис.
1.
Клетка E-coli с хромосомой и плазмидой.
Р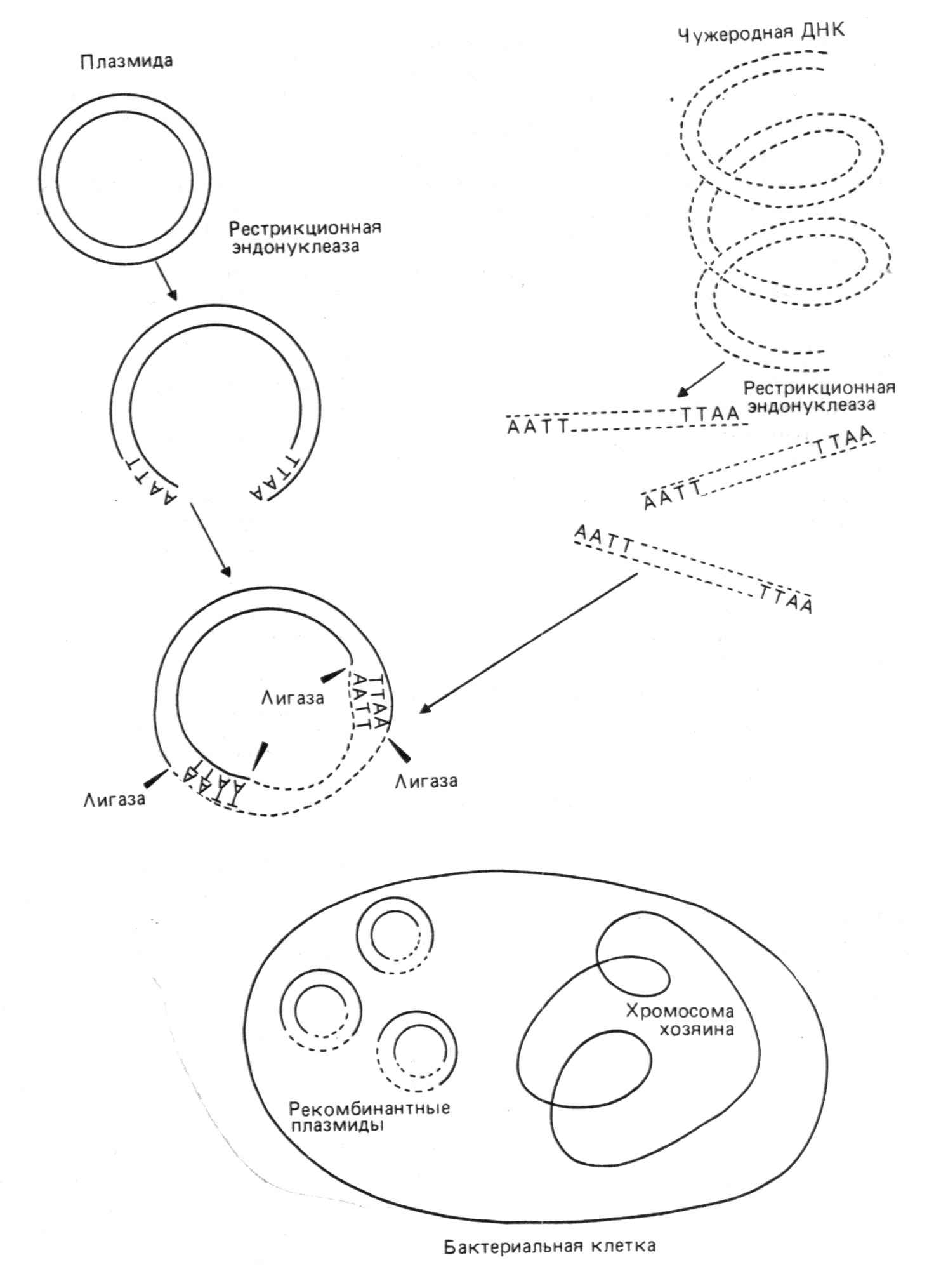
екомбинантную
конструкцию
вводят затем
в бактерию, где
она реплицируется
(см. рис.2 )
Рис. 2. Принцип введения чужеродной ДНК в бактериальную плазмиду с использованием эндонуклеазы.
Источник экзогенной ДНК не имеет значения. ДНК может быть получена, например, из клеток человека, но можно сшивать и искуственно синтизированные гены. Кроме бактериальных плазмид в качестве векторов (носителей) ДНК используют фаги λ (объект исследования Альберта). Часть генома этого фага не обязательна для его размножения в бактерии. Вместо него можно ввести чужеродную ДНК, которая будет размножаться вместе с фаговой, после инфицирования бактерий.
Добиться репликации и амплификации в составе плазмидной (или фаговой) ДНК после трансформации бактериальной клетки ещё не значит решить все её проблемы. Прежде всего возникают два вопроса:
Как распознавать клоны, содержащие гибридную ДНК, среди потомства трансформированных клеток или живых бактериофагов ?
как идентифицировать необходимые фрагменты ДНК среди многих клонированных неизвестных фрагментов?
Например можно отбирать бактериальные клетки, если они несут плазмиду с фактором устойчивости к антибиотику, выращивая их на среде, на среде, содержащей антибиотик. Нетрансформированные клетки без плазмид(и, следовательно, без гена устойчивости к антибиотику) просто не будут расти на такой среде. В последнее время разработано много специальных методов вакцинации, которые позволяют отбирать только рекомбинантные клетки.
Для генной инженерии белков недостаточно отобрать и размножить определённые фрагменты ДНК, необходимо ещё индуцировать их экспрессию в клетке. Для этого необходимо «подключить» рекомбинантную молекулу ДНК , последующую трансляцию матричной РНК и процессинг как на транскрипционном , так и на трансляционных уровнях.
1.4. ИДЕНТИФИКАЦИЯ И АНАЛИЗ ГЕНОВ.
Ещё одна область применения рестриктаз – идентификация и определение числа генов. Эти задачи решаются с помощью метода разработанного Саузерном.
Тотальную ДНК из клеток человека гидролизуют эндонуклеазой примерно на 500000 фрагментов длиной от 102 до 105 нуклеотидных пар. Затем фрагменты разделяют по молекулярной массе с помощью гель- электрофореза в ага розе, после чего ДНК денатурирует с щелочью прямо в геле, чтобы получить одноцепочные фрагменты. Их переносят на нитроцеллюлозный фильтр и фиксируют высушиванием при 800С. В результате получается отпечаток(реплика) картины разделения фрагментов ДНК по их размеру. Эти фрагменты можно идентифицировать методом гибридизации с радиоактивными ДНК-зондами, специфичными для определённых генов или хромосом. Любой фрагмент, содержащий всю последовательность зондируемого гена или его часть, будет выглядеть на радиоавтографе в виде тёмной полосы.
Зонды и генные библиотеки. Главное условие такого анализа - наличие подходящего геноспецифического радиоактивного ДНК-зонда, который можно использовать для гибридизации.
1.5. ГИБРИДИЗАЦИЯ НУКЛЕИНОВЫХ КИСЛОТ.
Способность к гибридизации цепей ДНК лежит в основе многих методических приёмов молекулярной биологии, поэтому более подробное описание принципа гибридизации будет полезным. Большинство природных ДНК встречается в виде двухцепочных молекул. Их устойчивость поддерживается благодаря тому, что пиримидиновое основание цитозин(C) спаривается с пуриновым основанием гуанином(G), в то время как пиримидиновое основание тимин(T) спаривается с пуриновым основанием аденином(A). Эти комплиментарные пары оснований удерживаются водородными связями(тремя в паре G-C и двумя в паре A-T), которые относительно легко разрываются, при этом одноцепочные фрагменты ДНК, присутствующие в растворе, снова формируют двойную спираль. Для реассоциации не имеет значения происхождения одноцепочной ДНК, не требуется даже полной комплиментарности отдельных цепей. Реассоциация происходит даже тогда, когда какая-то часть оснований в каждой цепи не комплиментарна. Одноцепочная ДНК может спариваться,то есть гибридизироваться даже с РНК, если у них есть комплиментарные основания.
"ПРОГУЛКА ПО ХРОМОСОМЕ". Метод гибридизации полезно
использовать, например, для анализа очень протяженного гена. При этом с помощью подходящего зонда из геномной библиотеки ДНК первоначально извлекается какая-то часть такого гена. Нуклеотидная последовательность этой части гена будет, как правило, длинее зонда, и её концы будут перекрываться с другими фрагментами данного гена в этой библиотеке,то есть будут, по крайней мере, частично гибридизироваться с ними. Свободные концы этих фрагментов будут гибридизироваться со следующими и так далее, пока весь структурный ген не будет полностью идентифицирован серией перекрывающихся фрагментов. Именно таким образом был реконструирован структурный ген фактора свертывания крови VII человека, необычно длинный, состоящий из 180000 пар нуклеотидов.
Существующий метод гибридизации in situ, этот метод уже усовершенствован на столько, что с его помощью можно локализовать в хромосомах даже уникальные гены, такие как ген инсулина. Вот пример некоторых генов человека, идентифицированных с помощью гибридизации(таблица 1).
ТАБЛИЦА 1.
Некоторые гены человека, идентифицированные с помощью гибридизации.
Ген, длина последовательности ДНК и число копий. |
Локализация. |
| ________________3__11 ρ | |
| ________________l__ 16 ρ | |
| Инеулин(900 п.н.) | 11 ρ 15 |
| Интерферон | 9 ρ 2,1 - pter |
| Онтоген fes(4000 п.н.) | 15 q 2,4 - gter |
| Сывороточный альбулин(1600 п.н.) | 4 q 11 - 22 |
| Миозин МНС(2200 п.н.) | 17 ρ1,2 - pter |
| Колаген(COL 1A2) | 7 q 22 |
1.6. СОРТИРОВКА ХРОМОСОМ.
Следующий метод – это метод сортировки хромосом при опмощи цитофлюрометрии. Этот метод может быть использован в двух разных целях:
1) Для идентификации и количественного анализа большого числа хромосом в течение очень короткого времени.
2) для препаративного разделения хромосом. Этот метод имеет два преимущества перед стандартными методами анализа хромосом:
во-первых, он полностью автоматический, благодаря чему исключается элемент субъективности
во-вторых, он намного быстрее (рис.3)
Однако важнее, что этот метод позволяет препаративно разделять хромосомы, и при наличии специфических зондов исследовать структуру и функцию отдельных генов становится относительно просто. В этом случае ген можно локализовать в хромосоме с помощью гибридизации in situ, размножить его ДНК путём клонирования и секвенирования.
Р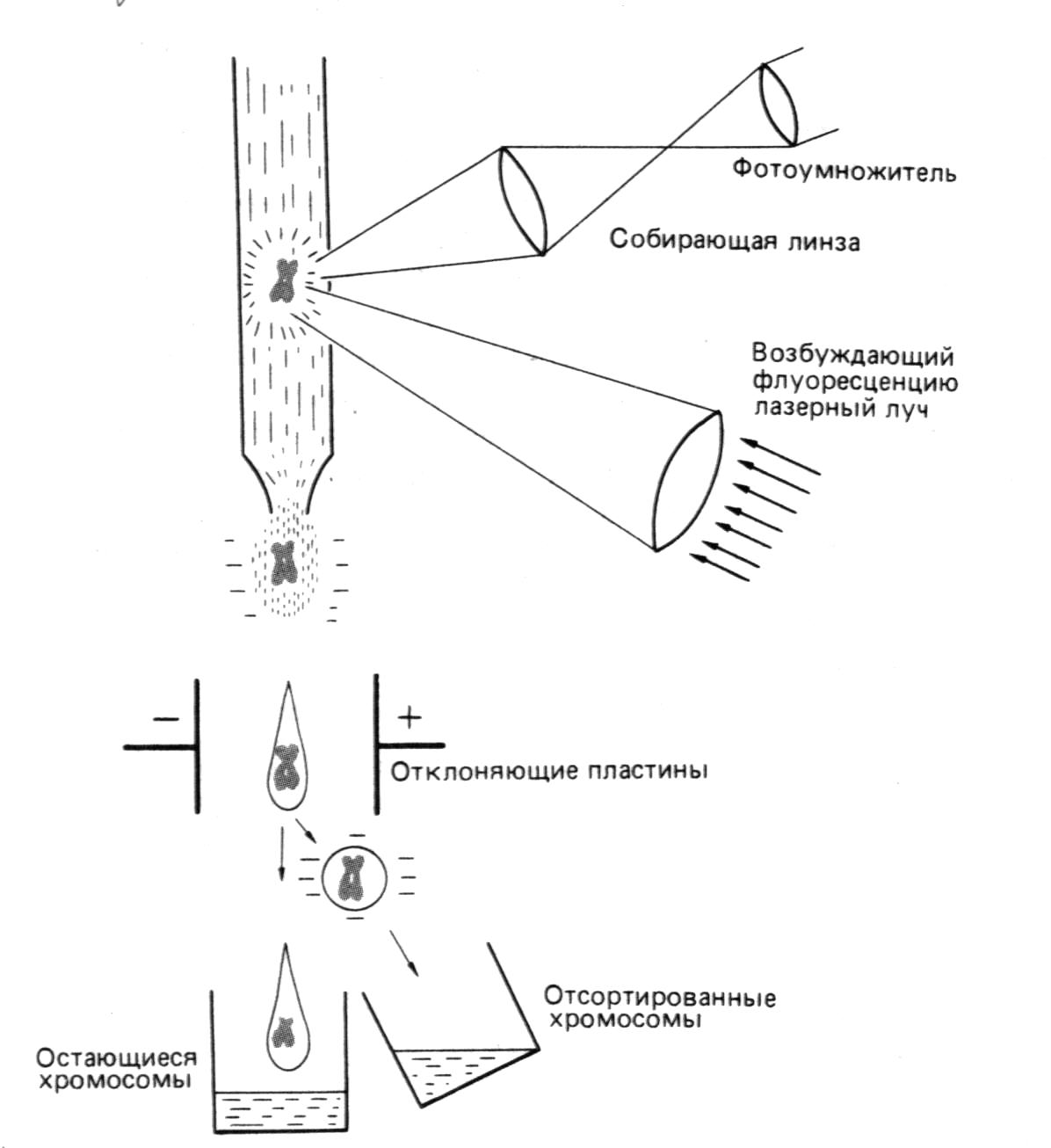
ис.
3. Принцип сортировки
хромосом с
помощью лазера.
Хромосомы
окрашены
флуоресцирующим
красителем.
Флуоресценция
возбуждается
лазерным лучом
и измеряется
для каждой
хромосомы
отдельно. Данные
измерений
используют
для сортировки
хромосом.
1.7. СЕКВЕНИРОВАНИЕ ДНК.
Последовательность нуклеотидов и генетический код.
Методы определённой последовательности аминокислот в полипептидной цепи были известны ещё в 50-х годах. Теоретически это относительно лёгкая проблема, поскольку все 20 аминокислот, встречающихся в природных белках, имеют разные свойства. С другой стороны, нуклеотидная последовательность ДНК относительно однородна по составу однородных звеньев, так как содержит только четыре типа азотистых оснований – гуанин, цитозин, аденин и тимин. Когда в 60-е годы был расшифрован генетический код, появилась возможность востанавливать (дедуцировать) нуклеотидную последовательность соответствующего белка. Однако генетический код является вырожденным, то есть одной и той же аминокислоте соответствуют несколько нуклеотидных триплетов. Следовательно сведения о нуклеотидной последовательности аминокислот в белке, не однозначны. Кроме того последовательности аминокислот не содержат никакой информации о последовательности некодирующих участков ДНК. Принцип состоит в следующем: длинную молекулу ДНК фрагментируют при помощи агентов, расщепляющихся в специфических сайтах. Затем определяют последовательность нуклеотидов в каждом из этих фрагментов. Очерёдность фрагментов в целой молекуле восстанавливают, используя перекрывающие концы: идентичные цепи разрезают повторно другой рестриктазой, а затем последовательности, перекрывающихся образующихся при обработке двумя рестриктазами разной специфичности, сравнивают. Так может быть реконструирована полная последовательность. В пределах отдельных фрагментов порядок нуклеотидов определяют с помощью специальных методов. Раньше секвенирование ДНК было весьма трудным делом, теперь же оно осуществляется очень легко и быстро. Для этого необходимо длинную молекулу ДНК с помощью рестриктазы разделить на фрагменты удобного размера, а затем, если нужно,мощью специальных методов. Раньше секвенирование ДНК было весьма трудным делом, теперь же оно осуществляется очень легко и быстро. Для этого необходимо длинную молекулу ДНК с помощью рестриктазы разделить на фрагменты удобного размера, а затем, если нужно,ные сведения и о нетранскрибирусных участках ДНК, важных для контроля транскрипции(так называемые операторы и промоторы).
1.8.ДИНАМИЧНОСТЬ ГЕНОМА.
Методы новой гентики расширили наши знания о структуре генетического материала. В 1963 году Тэйлор описал “индуцированные фагом мутации E. Coli”, вкоре после этого, Старлингер и Седлер описали IS-элементов у бактерий. Эти элементы получили название мобильных, теперь же они определяются как специфические последовательности ДНК, которые могут неоднократно внедряться в разные сайты генома. Перенос генов от одной бактерии к другой с помощью фага (трансдукция) известен давно, а теперь используется и в генетической инженерии эукариот (включая клетки млекопитающих). Возможно, такие процессы могут происходить и в природе. Более того, последовательности ДНК, гомологичные глобиновому гену человека, были обнаружены у бобовых растений. Функция такого гена у растений может состоять в том, чтобы “обеспечить кислородом клубеньковые бактерии в ткани”. Наличие этого гена может быть объяснено переносом его от насекомых или млекопитающих.(рис.4).
Стр 142 рис 2.95
НАРИСУЕМ САМИ
Итак, из выше изложенного можно сделать следующее заключение, что теоретическими предпосылками формирования генной инженерии как науки, явились:
Открытие двойной спирали ДНК.
Получение информации о матричном синтезе:
Репликации ДНК.
Транскрипции ДНК.
Трансляции ДНК.
3. Открытие плазмид.
4. Открытие фрагментов рестриктаз.
5. Осуществление процесса рекомбинации хромосом
Идентификация и анализ генов.
Способность к гибридизации цепей ДНК.
Секвенирование ДНК.
ГЛАВА 2. ВОЗМОЖНОСТИ ГЕННОЙ ИНЖЕНЕРИИ.
Значительный прогресс достигнут в практической области создания новых продуктов для медицинской промышленности и лечения болезней человека (табл.2).
ТАБЛИЦА 2.
Использование генно-инженерных продуктов в медицине.
|
Продукт |
Природные продукты и сфера применения генно-инженерных продуктов |
| Антикоагуляторы | Активатор тканевого плазминогена (АТП), активирует плазмин. Фермент, вовлечённый в рассасывание тромбов; эффективен при лечении больных инфарктом миокарда. |
| Факторы крови | Фактор VIII ускоряет образование сгустков; дефицитен у гемофиликов. Использование фактора VIII, полученного генно-инженерными методами, устраняет риск связанный с переливанием крови. |
| Факторы стимулирующие образование колоний | Ростовые факторы иммунной системы, которые стимулируют образование лейкоцитов. Применяют для лечения иммунодефицита и борьбе с инфекциями. |
| эритропоэтин | Стимулирует образование эритроцитов. Применяют для лечения анемии у больных с почечной недостаточностью. |
| Ростовые факторы |
Стимулируют дифференциацию и рост различных типов клеток. Применяют для ускорения лечения ран. |
| Гормон роста человека | Применяют при лечении карликовости. |
| Человеческий инсулин | Используется для лечения диабета |
| Интерферон | Препятствует размножению вирусов. Также используется для лечения некоторых форм раковых заболеваний. |
| Лейксины | Активируют и стимулируют работу различных типов лейкоцитов. Возможно применение при залечивании ран, при заражении ВИЧ, раковых заболеваний, иммунодефиците. |
| Моноклональные антитела | Высочайшая специфичность связанная с антителами используется в диагностических целях. применяют также для адресной доставки лекарств, токсинов, радиоактивных и изотопных соединений к раковым опухолям при терапии раков, имеется много других сфер применения. |
| Супероксид дисмутаз | Предотвращает поражение тканей реактивными оксипроизводными в условиях кратковременной нехватки кислорода, особенно в ходе хирургических операций, когда нужно внезапно восстановить ток крови. |
|
Вакцины |
Искуственно полученные вакцины (первой была получена вакцина против гепатита В) по многим показателям лучше обычных вакцин. |
В настоящее время фармацевтическая промышленность завоевала лидирующие позиции в мире, что нашло отражение не только в объёмах промышленного производства, но и в финансовых средствах, вкладываемых в эту промышленность (по оценкам экономистов, она вошла в лидирующую группу по объёму купли-продажи акций на рынках ценных бумаг). Важной новинкой стало и то, что фармацевтические компании включили в свою сферу выведение новых сортов сельскохозяйственных растений и животных, и тратят на это десятки миллионов долларов в год, они же мобилизировали выпуск химических веществ для быта. Добавок к продукции строительной индустрии и так далее. Уже не десятки тысяч, а возможно, несколько сот тысяч высококвалифицированных специалистов заняты в исследовательских и промышленных секторах фарминдустрии,и именно в этих областях интерес к геномным и генно-инженерным исследованиям исключительно высок.
Очевидно поэтому любой прогресс биотехнологий растений будет зависеть от разработки генетических систем и инструментов, которые позволят более эффективно управлять трансгенами. Ситуация аналогична той, которая наблюдается в компьютерной индустрии, где помимо увеличения объёмов обрабатываемой информации и улучшения самих компьютеров, нужны ещё и операционные системы управления информацией, типа микрософтовских “окон”.
Для чистого вырезания трансгенного ДНК в растительный геном, всё больше применяют заимствованные из микробной генетики системы гомологичной рекомбинации, такие как системы Cre-lox и Flp-frt. Будущее, очевидно, будет за управляемым переносом генов от сорта к сорту, основанного на применении предварительно подготовленного растительного материала, который уже содержит в нужных хромосомах участки гомологии, необходимого для гомологичного встраивания трангена. Помимо интегративных систем экспрессии, будут опробованы автономно реплицирующиеся векторы.осбый интерес представляют искуственные хромосомы растений, которые теоретически не накладывают никаких ограничений на объём вносимой теоретической информации.
Кроме этого учёные занимаются поиском генов, кодирующих новые полезные признаки. Ситуация в этой области меняется радикальным образом, прежде всего, существованию публичных баз данных, которые содержат информацию о большинстве генов, бактерий, дрожжей, человека и растений, а также в следствии разработки методов, позволяющих одновременно анализировать экспрессию большого количества генов с очень высокой пропускной способностью. Применяемые на практике методы можно разделить на две категории:
Методы, позволяющие вести экспрессионное профилирование: субстракционная гибридизация, электронное сравнение EST-библиотек, «генные чипы» и так далее. Они позволяют устанавливать корреляцию между тем или иным фенотипическим признаком и активностью конкретных генов.
Позиционное клонирование, заключается в создании за счет инсерционного мутагенеза мутантов с нарушениями в интересующем нас признаке или свойстве, с последующим клонированием соответствующего гена как такового, который заведомо содержит известную последовательность (инсерция).
Вышеназванные методы не предполагают ни каких изначальных сведений о генах, контролирующих тот или иной признак. Отсутствие рационального компонента в данном случае является положительным обстоятельством, поскольку неограничен нашими сегодняшними представлениями о природе и генном контроле конкретного интересующего нас признака.
Кроме всего этого группа ученых, таких как Марк Адам (ведущий сотрудник института геномных исследований в штате Мэриленд – США, частной исследовательской компании, занимающейся исключительной работой в области картирования генов), Крэйк Вентер (директор этого института) и соавторами, разрабатывается проект «Геном человека». Цель этого проекта заключается в выяснении последовательности оснований во всех молекулах ДНК в клетках человека. Одновременно должна быть установлена локализация всех генов, что помогло бы выяснить причину многих наследственных заболеваний и этим открыть пути к их лечению. Что бы последовательно приближаться к решению проблемы картирование генов человека, было сформулировано пять основных целей:
Завершить составление детальной генетической карты, на которой были бы помечены гены, отстоящие друг от друга на расстоянии не превышающем в среднем 2 млн. оснований (1 млн. оснований принято называть мегобазой);
составить физические карты каждой хромосомы (разрешение 0.1 Мб);
получить карту всего генома в виде охарактеризованных клонов (5 тыс. оснований в клоне или 5 Кб);
завершить к 2004 году полное секвенирование ДНК (разрешение одного основание);
нанести на полностью завершенную секвенсовую карту все гены человека (к 2005 году).
Ожидалось, что, когда все указанные цели будут постигнуты, исследователи определят все функции генов и разработают методы биологического и медицинского применения полученных данных.
Рассмотрев темпы ускорения работы в рамках проекта «Геном человека», руководители этого проекта объявили 23 октября 1998г., что программа будет полностью завершена гораздо раньше, чем планировалось, и сформулировали «Новые задачи проекта «Геном человека»:
полностью завершить в декабре 1998 года работу по секвенирование генома «Круглого червя» c. elegans (это было сделано в срок);
закончить предварительный анализ последовательности ДНК человека к 2001 году, а полную последовательность к 2003 году;
картировать к 2002 году геном плодовой мухи;
начать секвенирование генома мыши с использованием методов ДНК искусственных хромосом дрожжей (завершить этот проект к 2005 году).
Помимо этих целей, официально включен в поддерживаемый правительством США и рядом других правительств проект, некоторые исследовательские центры объявили о задачах, которые будут решаться в основном за счет частных фондов и пожертвователей. Так, ученые калифорнийского университета (Беркли), Орегонского университета и Ракового исследовательского центра имени Фрейда Хатчинсона начали программу «Геном собаки».
Международное общество секвенирование в феврале 1996 года приняло решение о том, что любая последовательность нуклиотидов размером 1-2 Кб должна быть обнародована (через Интернет) в течение 24 часов после ее установления.
2.1. ЧТО БУДЕТ СДЕЛАННО ПОСЛЕ ЗАВЕРШЕНИЯ АНАЛИЗА ГЕНОМА ЧЕЛОВЕКА.
Главная стратегическая задача будущего сформулирована следующим образом: изучить однонуклеотидные вариации ДНК в разных органах и клетках отдельных индивидуумов и выявить различия между индивидуумами. Анализ таких вариаций даст возможность не только подойти к созданию индивидуальных генных портретов людей, что в частности даст возможность лечить болезни, но и определить различия между популяциями. А также выявлять географические районы повышенного риска, что поможет давать чёткие рекомендации о необходимости очистке территории от загрязнения и выявить производства, на которых есть большая опасность поражение геномов персонала.
Эта грандиозная задача рождает не одни радужные ожидания всеобщего блага, но и вполне осознанную тревогу юристов и борцов за индивидуальные права человека. Так, в частности, высказываются возражения против распространения персональной информации без решения тех, кого она касается. Один пример помогает понять эти тревоги: уже сейчас страховые компании нацелились на добывание таких сведений правдами и неправдами, они намериваются использовать данные против тех, кого они страхуют. Например, если подающий на страховку несёт потенциально болезнетворный ген, компании не хотят страховать таких людей вовсе или же пытаются заломить бешенные суммы за их страховки. Исходя из этого, .конгресс США уже принял ряд законов, направленный на строгий запрет распространения генетической информации относительно отдельных людей, юристы всего мира интенсивно работают в данном направлении.
Глава 3. Области практического применения генной инженерии.
3.1. Создание трансгенных растений.
Еще 10 лет тому назад биотехнология растений заметно отставала в своем развитии, но за последние 3 года наблюдается быстрый выброс на рынок трансгенных растений с новыми полезными признаками. Трансгенные растения в США в 1996 году занимали площадь 3 млн. акров, в 1997 году площадь увеличилась до 15 млн. акров, в 1998 году – до 60 млн. акров, а в прошлом году до 80 млн. акров. Поскольку основные трансгенные формы кукурузы, сои, хлопчатника с устойчивостью к гербицидам и насекомым хорошо себя зарекомендовали, есть все основания ожидать, что площадь под генноиженерные растения в будущем (2001 году) увеличатся в 4-5 раз.
В апреле 1998 года доля в процентах трансгенных форм растений в сельском хозяйстве составило:
кукуруза – 6
соя – 12
хлопчатник – 15
томаты –
Так как число жителей за последнее столетие увеличилось с 1.5 до 5.5 млрд. человек, а к 2020 году предполагается вырост до 8 млрд., таким образом возникает огромная проблема, стоящая перед человечеством. Эта проблема заключается в огромном увеличение производства продуктов питания, несмотря на то, что за последние 40 лет производство увеличилось в 2.5 раза, все равно этого не достаточно. И в мире в связи с этим наблюдается социальный застой, который становится все более настоятельным. Другая проблема возникла с медицинским лечением. Несмотря на огромные достижение современной медицины, производимые сегодня лекарственные препараты столь дороги, что ѕ населения земли сейчас полностью полагаются на традиционные донаучные методы лечения, прежде всего на неочищенные препараты растительного происхождения.
В развитых странах лекарственные средства на 25% состоят из природных веществ, выделенных из растений. Открытия последних лет (противоопухолевые препараты: таксол, подофиллотоксин) свидетельствуют о том, что растения еще долго будут оставаться источником полезных биологически-активных веществ (БТА), и что способности растительной клетки к синтезу сложных БТА все еще значительно превосходят синтетические способности инженера-химика. Вот почему ученые взялись за проблему создания трансгенных растений.
Отсчёт истории генетической инженерии растений принято вести с 1982 года, когда впервые были получены генетически трансформированные растения. Метод трансформации основывается на природной способности бактерий Agrobacterium tumefaciens генетически модифицировать растения. Реконструированные штаммы Agrobactrium, содержащие неонкогенные варианты Ti-плазмид и обладающие повышенной вирулентностью, стали основой одного из наболее популярных методов трансформации. Первоначально трансформация применялась для генно-инженерных двудольных растений, однако работы последних лет свидетельствуют, что этот метод эффективен и в отношении кукурузы, риса, пшеницы. Другим широко распространённым методом трансформации, является технология, основанная на обстреле ткани микрочастицами золота (или других тяжелых металлов), покрытыми раствором ДНК. Все выращиваемые ныне коммерческие сорта получены с помощью названных выше двух методов.
Современный арсенал методов трансформации, однако, довольно обширен и включает такие подходы, как введение ДНК в голые клетки (протопласты), электропорация клеток, микроинъекций ДНК в клетки, прокалывание клеток путём встряхивания их в суспензии микроигл, опосредованная вирусами инфекции и так далее.
Генетические изменённые растения с устойчивостью к различным классам гербицидов в настоящее время являются наиболее успешным биотехнологическим продуктом. Дело в том, что биотехнология позволила совершить такой прыжок, так как оказалось возможным генетически изменять устойчивость растений к тем или иным гербицидам либо путем введения генов, кодирующих белки, нечувствительные к данному классу гербицидов, либо за счет введения генов, обеспечивающих ускоренный метаболизм гербицидов растений. К настоящему времени клонированы гены, кодирующие нечувствительные к действию гербицидов ферменты-мишени, что дало возможность получать трансгенные растения, устойчивые к таким гербицидам, как глифостат и хлорсульфуроновым, и имидазолиноновым гербицидом. Изолированы также гены, которые кодируют ферменты деградации некоторых гербицидов, что позволило получить трансгенные растения устойчивые к фосфинотрицину и далапону. В 1997 году устойчивая к Roundup соя, распространяемая компанией "As Grow", была признана в США сельскохозяйственным продуктом года.
Ученые пошли далее. Так как множество растений подвержены нападению и поеданию со стороны насекомых, то ученые генной инженерии провели эксперимент с давно известной бактерией Bacillus-Thiringiensis, которая продуцирует белок, оказалось она является очень токсичной для многих видов насекомых, но в то же время безопасна для млекопитающих., белок (дельта-эндотаксин, CRY-белок) продуцируется различными штамами Bacillus-Thiringiensis. Это прототаксин который расщепляется в кишечнике насекомых, образуя активированный токсин. Активизированный белок специфично связывается с рецепторами средней кешки насекомых, что приводит к образованию пор и лизису клеток кишечного эпителия. Взаимодействие токсинов с рецепторами строго специфично, что усложняет подбор комбинации токсин-насекомое. В природе найдено большое количество штаммов Bacillus-Thiringiensis, чьи токсины действуют только на определенные виды насекомых. Препараты Bacillus-Thiringiensis в течение десятилетий использовались для контроля насекомых на полях.
Встраивание гена этого белка в геном растений дает возможность получить трансгенные растения, не поедаемые насекомые. Но этот метод потребовал большой работы со стороны генной инженерии, в плане подборов необходимых штаммов и созданию генно-инженерных конструкций, которые дают наибольший эффект для конкретных классов насекомых. Кроме видоспецифичности по действию на насекомых встраивание прокариотических генов дельта-токсинов в геном растений даже под контролем сильных эукариотических промоторов не привело к высокому уровню экспрессии. Предположительно такое явление возникло в связи с тем, что эти бактериальные гены содержат значительно больше адениновых и тиминовых нуклеатидных оснований, чем растительная ДНК. Эта проблдема была решена путем создания модефицированных генов, где один из природного гена вырезали и добавили те или иные фрагменты с сохранением доменов, кодирующих активные части дельта-токсинов. Так, например, с помощью таких подходов был получен картофель, устойчивый к колорадскому жуку. В настоящее время так называемый Bt – растения хлопка и кукурузы занимают основную долю в общем объеме генетически модифицированных растений этих культур, которые выращивают на полях США.
3.1.1 Изменение свойств сельскохозяйственных технических растений
Современная биотехнология в состоянии манипулировать многими важнейшими признаками, которые можно разделить на три группы:
Сельскохозяйственные производства. К ним можно отнести общей продуктивности растений за счет регулирования синтеза фитогормонов или дополнительного снабжения кислородом растительных клеток, а также признаки обеспечивающие устойчивость к разного рода вредителям, кроме этого в создании форм растений с мужской стерильностью и возможностью дольше сберегать урожай.
К признакам которые влияют на качество продукции, относится возможность манипулировать молекулярным весом жирных кислот. Растения будут производить биодеградирующий пластик, по цене сопоставимой с полиэтиленом, получаемым из нефти. Открылась возможность получения крахмала с заданными физико-химическими свойствами. Аминокислотный состав у растений запасных белков становится более сбалансированным и легко усвояем для млекопитающих. Растения становятся продуцентами вакцин, фармакологических белков и антител, что позволяет удешевить увеличение разных заболеваний, в том числе и онкологических. Получены и испытываются трансгенные растения хлопка с уже окрашенным волокном, более высоким качеством.
3.1.2. Генетическая модификация пластид.
Во многих случаях генетической модификации будут подвергаться не ядерные геномы, а геномопластит или метохондрия. Такие системы позволяю значительно увеличить содержание продукта в трансгенном материале.
В генной инженерии исследуются следующие направления:
Управляемая активность генов;
Селективная экспрессия трансгена в определенных тканях;
Система экспрессии растения в чужеродной генетической информации, опосредованной вирусами.
Разработанная усилиями компании “Biosource” (США) технология позволяет быстро и в больших количествах нарабатывать в растениях белки и небольшие молекулы за счет инфицирования растений генетически модифицированными вирусами, со встроенными чужеродными генами тех или иных белков. За этой системой большое будущее так как она позволяет изменить биосинтетические процессы в растениях без длительных и дорогостоящих манипуляций с растительным геномом.
3.3. ГЕННЫЕ ВАКЦИНЫ
3.2.1. Актуальность разработки новых вакцин
Вакцины — одно из самых значительных достижений медицины, их использование к тому же чрезвычайно эффективно с экономической точки зрения. В последние годы разработке вакцин стали уделять особое внимание. Это обусловлено тем, что до настоящего времени не удалось получить высокоэффективные вакцины для предупреждения многих распространенных или опасных инфекционных заболеваний. По данным созданной в прошлом году международной организации «Всемирный союз по вакцинам и иммунизации» (в числе ее участников — ВОЗ, ЮНИСЕФ, Международная федерация ассоциаций производителей фармацевтической продукции, Программа Билла и Мелинды Гейтс по вакцинации детей, Рокфеллеровский фонд и др.), в настоящее время отсутствуют эффективные вакцины, способные предупредить развитие СПИДа, туберкулеза и малярии, от которых в 1998 г. умерло около 5 млн человек. Кроме того, увеличилась заболеваемость, обусловленная теми инфекциями, с которыми человечество ранее успешно боролось. Этому способствовало появление лекарственно-устойчивых форм микроорганизмов, увеличение числа ВИЧ-инфицированных пациентов с иммунной недостаточностью, ослабление систем здравоохранения в странах с переходной экономикой, увеличение миграции населения, региональные конфликты и др. При этом распространение микроорганизмов, устойчивых к воздействию антибактериальных препаратов, приобрело характер экологической катастрофы и поставило под угрозу эффективность лечения многих тяжелых заболеваний. Повышенный интерес к вакцинам возник после того, как была установлена роль патогенных микроорганизмов в развитии тех заболеваний, которые ранее не считали инфекционными. Например, гастриты, пептическая язва желудка и двенадцатиперстной кишки, ассоциированная с H. pylori, злокачественные новообразования печени (вирусы гепатита В и С).
Поэтому в последние 10–15 лет правительства многих стран стали принимать меры, направленные на интенсивную разработку и производство принципиально новых вакцин. Например, в США в 1986 г. был принят закон («National Vaccine Injury Compensation Act»), защищающий производителей вакцин от юридической ответственности при подаче судебных исков, связанных с развитием побочных реакций при вакцинации, если они не были обусловлены ошибками при производстве вакцины. С изменением ситуации увеличился и мировой рынок вакцин, объем продаж которого в 1998 г. составил 4 млрд долларов США в стоимостном выражении. Однако многие считают, что в ближайшие годы этот сектор фармацевтической промышленности будет развиваться гораздо быстрее. Так, согласно публикациям в американском журнале «Signals Magazine» (январь 1999 г.), который освещает ситуацию в современной биотехнологической промышленности, объем продаж вакцин на мировом рынке через 10 лет составит 20 млрд долларов США. Этот прогноз принадлежит М. Греко, исполнительному директору компании «Merieux MSD», совместного предприятия крупнейших производителей вакцин — компаний «Pasteur Merieux Connaught» (теперь «Aventis Pasteur») и «Merck & Co.».
3.2.2.Разработка ДНК-вакцин
Используемые сегодня вакцины можно разделить в зависимости от методов их получения на следующие типы:
• живые аттенуированные вакцины;
• инактивированные вакцины;
• вакцины,
содержащие
очищенные
компоненты
микроорганизмов
(протеины или
полисахариды);
• рекомбинантные
вакцины, содержащие
компоненты
микроорганизмов,
полученные
методом генной
инженерии.
Технологию рекомбинантной ДНК применяют также для создания живых ослабленных вакцин нового типа, достигая аттенуации путем направленных мутаций генов, кодирующих вирулентные протеины возбудителя заболевания. Эту же технологию используют и для получения живых рекомбинантных вакцин, встраивая гены, кодирующие иммуногенные протеины, в живые непатогенные вирусы или бактерии (векторы), которые и вводят человеку.
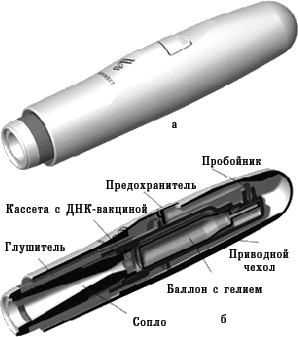
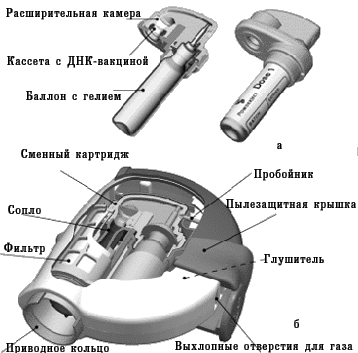
В 1990 г. в некоторых исследовательских лабораториях приступили к разработке новых вакцин, которые основаны на введении «голой» молекулы ДНК. Уже в 1992–1993 гг. несколько независимых групп исследователей в результате эксперимента доказали, что введение чужеродной ДНК в организм животного способствует формированию иммунитета. Принцип применения ДНК-вакцин заключается в том, что в организм пациента вводят молекулу ДНК, содержащую гены, кодирующие иммуногенные белки патогенного микроорганизма. ДНК-вакцины называют еще генными, генетическими, полинуклеотидными вакцинами, вакцинами из нуклеиновых кислот. На совещании специалистов по генным вакцинам, проведенном в 1994 г. под эгидой ВОЗ, было решено отдать предпочтение термину «вакцины из нуклеиновых кислот» с их подразделением соответственно на ДНК- и РНК-вакцины. Такое решение основывалось на том, что употребление термина «ДНК-вакцина» не сформирует ошибочное мнение о том, что новые вакцины вносят изменения в генетические структуры организма вакцинируемого человека. Тем не менее, многие специалисты считают более точным термин «генные вакцины» (поскольку иммунная реакция направлена не против ДНК, а против антигенного белка, кодируемого геном), который также часто применяют. Для получения ДНК-вакцин ген, кодирующий продукцию иммуногенного протеина какого-либо микроорганизма, встраивают в бактериальную плазмиду. Плазмида представляет собой небольшую стабильную молекулу кольцевой двухцепочечной ДНК, которая способна к репликации (воспроизведению) в бактериальной клетке. Кроме гена, кодирующего вакцинирующий протеин, в плазмиду встраивают генетические элементы, которые необходимы для экспрессии («включения») этого гена в клетках эукариотов, в том числе человека, для обеспечения синтеза белка. Такую плазмиду вводят в культуру бактериальных клеток, чтобы получить большое количество копий. Затем плазмидную ДНК выделяют из бактерий, очищают от других молекул ДНК и примесей. Очищенная молекула ДНК и служит вакциной. Введение ДНК-вакцины обеспечивает синтез чужеродных протеинов клетками вакцинируемого организма, что приводит к последующей выработке иммунитета против соответствующего возбудителя. При этом плазмиды, содержащие соответствующий ген, не встраиваются в ДНК хромосом человека. ДНК-вакцины можно вводить в солевом растворе обычным парентеральным способом (внутримышечно, внутрикожно). При этом бoльшая часть ДНК поступает в межклеточное пространство и только после этого включается в клетки. Применяют и другой метод введения, используя так называемый генный пистолет (рис. 5, 6). Для этого ДНК фиксируют на микроскопических золотых гранулах (около 1–2 мкм), затем с помощью устройства, приводимого в действие сжатым гелием, гранулы «выстреливают» непосредственно внутрь клеток. Следует отметить, что аналогичный принцип введения лекарства с помощью струи сжатого гелия используют и для разработки новых способов доставки лекарственных средств (с этой целью оптимизируют размеры частиц лекарственного вещества и их плотность для достижения необходимой глубины проникновения в соответствующую ткань организма). Этот метод требует очень небольшого количества ДНК для иммунизации. Если при иммунизации классическими субъединичными вакцинами вводят микрограммы протеина, то при использовании ДНК-вакцины — нанограммы и даже меньше. Говоря о минимальном количестве ДНК, достаточном для индукции иммунного ответа, С.А. Джонстон, директор Центра биомедицинских изобретений Техасского университета, в журнале «The Scientist» (1998) отмечает, что с помощью генного пистолета можно однократно ввести мыши «фактически 27 тыс. различных плазмид и получить иммунный ответ на индивидуальную плазмиду».
|
|||||||||||||||||||||||||||||||||||||||||||||||||||