

 Реферат: Биохимия нуклеиновых кислот
Реферат: Биохимия нуклеиновых кислот
Реферат: Биохимия нуклеиновых кислот
Д. Г. Кнорре
Новосибирский государственный университет
Информационное содержание ДНК. Регуляторные районы и интроны
В предыдущей статье1 было рассказано лишь о самом главном информационном содержании ДНК: ДНК содержит программы для синтеза свойственных данному организму белков, которые представляют собой последовательности троек нуклеотидов на транскрибируемой нити. Непосредственно они программируют синтез соответствующего кодона мРНК. Например, будущий остаток фенилаланина должен быть записан на транскрибируемой нити ДНК как (3')dAdAdA(5'), если программируется ко-дон UUU, или как (3')dAdAdG(5'), если кодируется кодон UUC. Участок ДНК, содержащий всю информацию о программируемом белке, называют геном для данного белка. Генами называют и участки ДНК, которые содержат всю необходимую информацию о структуре РНК (рибосомные и транспортные РНК), а также вспомогательные участки, необходимые для транскрипции гена. В действительности последовательностью кодирующих три-нуклеотидов для синтеза белков или последовательностью нуклеотидов, кодирующих структуру тРНК или рРНК, содержащаяся в ДНК информация не исчерпывается. Сегодня уже ясно, что оно неизмеримо богаче, а, по-видимому, многое еще и вообще неизвестно.
Для создания определенной РНК необходимо, чтобы транскрипция началась в определенной точке огромной молекулы ДНК. Здесь должна оказаться и расположиться нужным для транскрипции образом РНК-полимераза. Для этого существует специальная область, чаще всего находящаяся перед кодирующей последовательностью, роль которой состоит в том, чтобы связать и ориентировать определенным образом РНК-полимеразу. Эту область, если она расположена вдоль цепи близко от начала старта транскрипции, называют промотором.
Новое неожиданное оказалось связанным со структурой генов эукариот. Выяснилось, что смысловая последовательность, кодирующая определенную последовательность аминокислот в белке или определенную последовательность нуклеотидов в РНК, необязательно является непрерывной. Она может содержать некоторые вставочные последовательности, которые получили название интронов.
1 Кнорре Д. Г. Биохимия нуклеиновых кислот // Соросов -ский Образовательный Журнал. 1996. № 3. С. 11—16.
Кодирующие последовательности, разделенные интронами, в этом случае называют экзонами. При транскрипции получаются РНК-копии, которые содержат и нужные в конечной (зрелой) молекуле РНК фрагменты, и лишние, которые должны быть удалены (вырезаны) из зрелой молекулы. Каждое такое вырезание должно сопровождаться воссоединением концов последовательных экзонов, между которыми находился интрон. Этот процесс получил название "сплайсинг". Сплайсинг происходит с избирательностью, характерной для процессов, катализируемых ферментами. До открытия сплайсинга считалось общепринятым, что все ферменты имеют белковую природу. Однако никакие белки в некоторых случаях в сплайсинге не участвуют, то есть РНК, содержащая интроны, может сама осуществлять селективную реакцию, причем с довольно большой скоростью. В результате пришлось признать, что биологическими катализаторами могут служить не только белки, но и рибонуклеиновые кислоты. По аналогии с энзимами такие катализаторы получили название рибозимов.
Говоря об информационном содержании ДНК, нельзя не сказать о том, что информационное содержание ДНК может изменяться как в результате ошибок при репликации, так и при действии различных внешних факторов — облучения и обработки некоторыми химическими реагентами. Изменения в структуре ДНК какого-либо организма носят название мутаций. Мутация может заключаться в том, что одна пара нуклеотидов в двух цепях превращается в другую комплементарную пару. Это, естественно, происходит не единовременно: сначала возникает одна ошибка, например dC заменяется на dT. При репликации dT отберет для введения в новую цепь dA, а не dG, которое находилось в этом месте цепи у родительской ДНК. В итоге вместо пары dC • dG в дочерней молекуле, а тем самым и во всех последующих поколениях на этом месте будет находится пара dT • dA. Такие мутации получили название точечных. Происходят и более значительные по масштабу мутации. Часто, хотя далеко не всегда, мутации имеют серьезные биологические последствия. Известны, например, гены, которые отвечают за контроль над размножением клеток, стимулируя его до определенного предела и останавливая в нужной для организма фазе. Некоторые единичные изменения в таком гене приводят к тому, что способность к контролю за процессом деления клеток теряется и ген превращается в онкоген, то есть ген, способствующий неограниченному размножению клеток — клетки становятся злокачественными и возникает раковая опухоль.
Что современная химия и биохимия умеют делать с нуклеиновыми кислотами
Химия и биохимия нуклеиновых кислот не только углубили наши представления об огромной группе важнейших биологических процессов, связанных с сохранением, размножением и использованием наследственной информации, заложенной в генетическом материале клеток, но дали мощный импульс для становления современной биотехнологии с важными практическими выходами. Для того чтобы не вслепую манипулировать нуклеиновыми кислотами, было необходимо научиться изучать их структуру, прежде всего составляющие их последовательности нуклеотидов. Впервые ценой воистину героических усилий это было сделано для нескольких тРНК, состоящих из нескольких десятков нуклеотидов. Приятно отметить, что на первом этапе соревнования за установление первичной структуры тРНК в лидирующей пятерке была и группа советских ученых, возглавляемая А.А. Баевым. Однако созданные в ходе этих работ методы были настолько трудоемки, что для прорыва в исследование ДНК и больших молекул РНК они были непригодны. Революционные решения, принципиально отличающиеся как друг от друга, так и от первых методов, были найдены в США Алленом Максамом и Уолтером Гилбертом и в Англии Фредериком Сенгером. Эти методы, в особенности метод Сенгера, сделали возможным секвенирование нуклеиновых кислот общим размером в миллионы нуклеотидных пар.
Конечно, в рамках одной статьи рассказать в деталях об этих методах невозможно. Поэтому придется ограничиться изложением наиболее принципиальных особенностей метода, причем только метода Сенгера. Особенностью проблемы является необходимость определить последовательное расположение очень большого числа остатков нуклеотидов (за один проход — несколько сот). Зато свойства каждого остатка уже хорошо изучены. Основополагающая идея Сенгера состояла в том, чтобы исследовать последовательность не самой ДНК или, точнее, ее достаточно длинного фрагмента, а исследовать структуру новосинтезированной ДНК, полученной на исследуемой ДНК как матрице с помощью ДНК-полимеразы. Из-за комплементарности новой ДНК по ее последовательности нуклеотидов нетрудно восстановить структуру матрицы. При этом оказалось возможным частично заменить в смеси мономеров, из которой синтезируется новая цепь, один из них на так называемый дидезоксинуклеотид. На рис. 1 приведена химическая формула нуклеотида, а именно тимидинфосфата, обозначенного символом dT. Остатком фосфорной кислоты он связан в цепи с предыдущим нуклеотидом. Его ОН-группа должна присоединить следующий нуклеотид при продолжении роста цепи. Оказалось, что при репликации можно совершить небольшой обман ДНК-полиме-разы — подмешать к смеси нуклеотидов дидезокси-
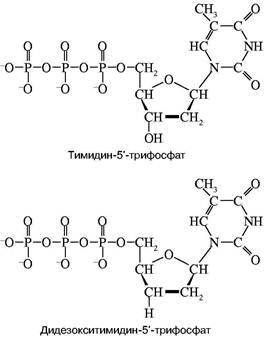
Рис. 1. Структуры тимидин-5'-трифосфата и его дидезоксианалога
рибонуклеотид, у которого эта ОН-группа отсутствует. Его можно обозначить в представленном случае символом ddT. С какой-то вероятностью ДНК-полимераза узнает и присоединит к растущей цепи очень похожий ddT вместо dT. Так как синтез идет в строгом соответствии с принципом компле-ментарности, то это произойдет напротив той точки матричной ДНК, где находится комплементарный dA. Но из-за отсутствия у ddT ОН-группы эта цепочка не сможет расти дальше, произойдет обрыв цепи. Обрыв с определенной вероятностью может произойти в любой точке напротив dA. Таким образом, получится смесь новосинтезированных цепей разной протяженности, причем длины (числа нук-леотидных остатков) всех этих цепей точно соответствуют номерам остатков dA матрицы. Следовательно, в таком эксперименте определяются точки расположения всех остатков dA в исследуемой ДНК. Если три аналогичных эксперимента провести со смесями, содержащими примеси других диде-зоксинуклеотидов: ddA, ddC и ddG, то аналогично будут расставлены по номерам все остальные три нуклеотида на исследуемой ДНК. Разделить же полученные смеси по длинам не представляет труда с помощью электрофореза — метода, основанного на перемещении заряженных молекул под действием постоянного электрического поля. Дело в том, что каждый остаток нуклеотида содержит остаток фосфорной кислоты, который несет отрицательный заряд. Поэтому, будучи помещены в электрическое поле, фрагменты разной длины будут перемещаться к аноду с разными скоростями. Так как электрофорез обычно проводят в очень вязкой среде (геле), то эта среда оказывает сопротивление перемещению фрагментов, тем большее, чем больше размером фрагмент. Этот фактор пересиливает действие поля, поэтому, чем длиннее фрагмент, тем медленнее он двигается, но все они располагаются в порядке, соответствующем их длинам. Остается только "увидеть" место каждого фрагмента. До сих пор для этой цели чаще всего используют радиоактивную метку: в нуклеотиды, из которых синтезируется новая цепь, вводят радиоактивный фосфор. Поэтому после окончания гель-электрофореза гель прикладывают к рентгеновской пленке и на месте нахождения фрагментов после проявления радиоавтографа появляются темные пятна. На рис. 2 схематично показаны такой радиоавтограф и читаемая с него последовательность маленького кусочка ДНК.
Страницы: 1, 2
