

 Курсовая работа: Определение активности ферментов
Курсовая работа: Определение активности ферментов
Субстратом служит 2%-ный раствор гемоглобина в 0.06 н. растворе соляной кислоты, 5 мл субстрата нагревают до 250С, прибавляют 1 мл раствора фермента, оставляют в течении 10 мин., добавляют 10 мл 0.3 н. раствора трихлоруксусной кислоты, энергично взбалтывают, фильтруют и 5 мл фильтрата отливают в мерную колбу на 25 мл. Сюда же приливают 10 мл 0.5 н. раствора едкого натра и 3 мл фенольного реактива (реактив Фолина-Чиокалтеу разводят предварительно двойным объемом воды) и доводят водой до метки. По истечении нескольких минут окрашенный раствор фотометрируют относительно стандартного раствора, содержащего 0.0008 мэкв тирозина в 5 мл 0.2 н. раствора соляной кислоты (к раствору добавляют 0.5% формальдегида в качестве антисептика). Для исключения возможных ошибок ставят холостой опыт. Единицы пепсина выражают в миллиэквивалентах тирозина (в пределах от 6.10-4 до 11.10-4): единицы пепсина = миллиэквиваленты тирозина х1.47.
Определение протеолитических ферментов по Муру и Штейну. Метод основан на том, что содержащие a-аминогруппу аминокислоты дают с нингидридом окрашенную производную - дикетогидриндилидендикетогидриндамин, а также альдегид-аминокислоты и углекислоту.
Окрашенный продукт обладает характерным максимумом поглощения при 570 мm, а интенсивность окраски зависит от количества аминокислоты. С пролином и оксипиролином реакция протекает в ином направлении, а максимум поглощения получаемого при этом окрашенного продукта расположен при 440 мm.
Инкубацию фермента удобно проводить в мерных колбочках по 5 мл, в которых смешивают и уравновешивают при постоянной температуре на водяной бане все компоненты за исключением фермента. После этого добавляют фермент, раствор смешивают и берут пробы для исследования. Для кинетических измерений концентрацию фермента желательно подобрать таким образом, чтобы аликвотные части можно было бы брать каждые 5 минут.
Аликвотные части пробы, взятые сразу же после добавления фермента, а затем через соответствующие интервалы, добавляют в фотометрические пробирки, содержащие 2 мл реактива нингидрида - это тотчас обрывает реакцию. После этого смесь 20 мин. нагревают на водяной бане; полученная окраска устойчива минимум 24 часа. Средняя ошибка при анализе повторных проб составляет ±2%. После разбавления кипяченой смеси пробы и нингидрида 10 мл смеси, состоящей из равных объемов воды и н.пропанола, интенсивность окраски определяют на спектрофотометре Колемана при 570 мm.
Для калибровки пользуются постоянными количествами стандартных аминокислот, содержащих реакционные компоненты. При помощи калибровочных кривых, построенных отдельно для каждой исследуемой аминокислоты, измерения интенсивности окраски (по отношению к контрольной колбе) пересчитывают в миллимоли. При делении этих величин на миллимоли субстрата, использованного в процессе реакции, можно найти процент гидролиза.
Построение калибровочной кривой исключает ошибки, происходящие от качественных различий реакции аминокислот на цветной реактив. До оптической плотности 0.1 зависимость между величиной поглощения и количеством субстрата носит линейный характер.
Воспроизводимость составляет ±3%. Мешающее влияние аммония, аминов и других веществ, содержащихся в биологическом материале и дающих цветную реакцию с нингидридом, исключается при помощи соответствующих холостых проб. Анализ сравнительно упрощается, когда изучают протеолитические ферменты, субстратами которых являются пептиды, не содержащие свободных аминогрупп. В этом случае вся образующаяся окраска целиком приходится на освободившуюся аминокислоту.
Микрометод определения активности протеиназ А.П.Алексеенко. Предлагаемый микрометод определения активности протеолитических ферментов основан на измерении содержания аргинина в пептидах, освобождающихся при гидролизе протеиназами белковых субстратов. Содержание аргинина определяют с помощью стабилизированной и усовершенствованной реакции Сакагучи (хроматограмму погружают в 0.1% раствор 8-гидроксихинолина в ацетоне, высушивают на воздухе, опрыскивают раствором 0.2 мл Br2 в 100 мл 0.5 М NaOH; аргинин и другие гуанидины дают оранжево-красные пятна, тауромицин и гликоциамин - лишь временную окраску), позволяющей определить содержание аргинина в растворах трихлоруксусной кислоты после осаждения и удаления нерастворимых белков. Зная содержание аргинина в белковом субстрате и определяя его количество в перешедших в раствор пептидах, можно рассчитать процент гидролиза белкового субстрата изучаемыми протеиназами. В этом состоит главное преимущество метода, поскольку метод Ансона не дает возможности рассчитать процент гидролиза белка, атакуемого протеолитическими ферментами. Из всех существующих методов только метод Мура и Штейна позволяет определить процент гидролиза белковых субстратов, но он имеет ограниченное применение, поскольку требует предварительного проведения полного кислотного гидролиза как белка-субстрата, так и образовавшихся пептидов. В предложенной методике калибровочная кривая строится по растворам свободного аргинина. Введение в реакцию пептидов и белков в виде их биуретовых комплексов повысило чувствительность реакции Сакагучи с остатками аргинина и сделало ее такой же чувствительной, как и для свободного аргинина. В качестве белкового субстрата используют классический субстрат - гемоглобин и окисленный по Сангеру лизоцим. Этот низкомолекулярный белок, содержащий 12.7% аргинина прекрасно расщепляется пепсином (рвется около 30 связей).
Прежде чем приступить к определению активности ферментов в изучаемых тканевых субстратах, необходимо:
1. Определить зависимость активности от рН и выбрать оптимальную реакцию среды для проявления активности изучаемого фермента.
2. Построить график зависимости активности фермента от времени его инкубации с субстратом. выбрать для работы те сроки инкубации, при которых сохраняется линейная зависимость между величиной активности фермента и временем его инкубации.
3. Определить зависимость величины активности от концентрации белка в пробе. Выбрать такие концентрации фермента, при которых величина активности фермента была бы пропорциональна его концентрации. Следует учитывать, что в тканевых экстрактах и биологических жидкостях могут присутствовать ингибиторы протеолитических ферментов. При разведении пробы их концентрация снижается , а протеолитических - увеличивается. Определяя “фактор разведения”, можно одновременно с подбором оптимальных условий для определения протеолитической активности выявить и наличие ингибитора.
4. При определении активности фермента необходимо работать при постоянной скорости ферментативной реакции, достигаемой при полном насыщении фермента субстратом, при так называемой максимальной скорости ферментативной реакции. В каждом отдельном случае максимальную скорость необходимо найти экспериментально, измерив активность препарата при разных концентрациях субстрата, при постоянных концентрациях белка и времени инкубирования.
Определение активности протеиназ по белковому субстрату.
К 0.5 мл субстрата в соответствующем буферном растворе добавляют 0.5 мл пробы, содержащей фермент (экстракт, биологическую жидкость), инкубируют установленное опытным путем время , после чего белки в пробе осаждают 5 мл 10% трихлоруксусной кислотой. Отделяют осадок центрифугированием, а надосадочную жидкость (ТХУ-центрифугат) подвергают дальнейшей обработке. Контрольные пробы - пробы, где реактив добавлен в обратном порядке: к 3 мл 10% раствора ТХУ добавляют 0.5 мл раствора, содержащего фермент и 0.5 мл субстрата. Рекомендуется ставить пробу на автолиз субстрата: к 0.5 мл субстрата добавляют 0.5 мл прокипяченного раствора, содержащего фермент, инкубируют вместе с опытными пробами и осажденной ТХУ.
Определение содержания аргинина в ТХУ-центрифугантах по модифицированной реакции Сакагучи. К 0.5 мл ТХУ-центрифуганта добавляют 0.5 мл 2.5мМ раствора CuSO4 , 0.5 мл 5 Н. КОН, 0.5 мл раствора ДХН (2,4-дихлор-1-нафтол). Раствор встряхивают и вносят 0.5 мл гипобромита натрия, мгновенно возникает ярко-розовая окраска. Раствор встряхивают и через 20-30 секунд стабилизируют пробы добавлением 0.2 мл раствора 2-тиогликоля (чтобы предотвратить разрушение окрашенного продукта избытком гипобромита натрия. После добавления 2-тиодигликоля в пробы как опытные, так и контрольные, появляется желтоватое окрашивание за счет побочного продукта реакции между избыточной ТХУ и гипобромита натрия. Побочный продукт также стабилизируется 2-тиодигликолем. Оптическая плотность пробы измеряется спектрофотометром при 520 нм в кювете толщиной 1 см.
Определение активности протеиназ по реакции с реактивом Фолина. К 0.5 мл того же ТХУ-центрифуганта добавляют 0.5 мл 2.5 мМ раствора CuSO4 , 4 мл 0.5 н. раствора NaOH и 1.5 мл разбавленного в три раза реактива Фолина. Через 30 минут измеряют на спектрофотометре оптическую плотность при 760 нм
Калибровочные кривые по пепсиновому гидролизату окисленного лизоцима. 30 мг окисленного лизоцима растворяют в 50 мл подкисленной воды (40 мл Н2О + 10 мл 0.3 н. раствора HСl. и к полученном раствору добавляют 0.5 мг пепсина. Смесь оставляют при комнатной температуре на сутки. По истечении этого срока инкубации препарат не дает осадка с ТХУ. Этот препарат используют наряду с растворами аргинина и тирозина в качестве стандарта для построения калибровочных кривых. Приготовляют разведения с содержанием от 60 до 600 мкг/мл исходного лизоцима. В пробы вводят соответствующее опытным пробам количество ТХУ. Приготовляют также растворы свободного аргинина и тирозина с концентрациями от 0.04 до 0.25 мкмоль/мл. Из каждой пробы берут по 0.5 мл (в трех параллельных пробах) для определения содержания аргинина и тирозина по Фолину.
Калибровочные кривые. 3 получена с помощью микрометода на основе реакции Сакагучи (520 нм) по растворам аргинина (х) и по пептидному гидролизу лизоцима (о); 1 и 2 - с помощью реакции Фолина (760 нм): 2 - по растворам тирозина, 1 - по пептидному гидролизу лизоцима. Значение оптической плотности
проб, измеренные при 520 нм или 760 нм, нанесены на
график против концентрации аргинина или тирозина или их остатков в пепсиновом
гидролизате лизоцима. Кривая 1, проходящая выше кривой 2, получена при реакции
реактива Фолина с рас-
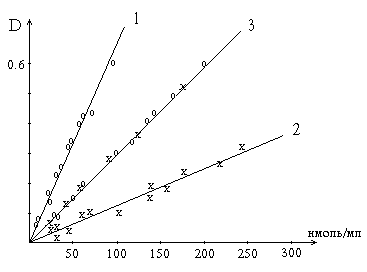
Количество аминокислоты или ее остатка
в гидролизате в наномолях на 1 мл.
творами тирозина эквивалентной концентрации. Кривая 3 получена в результате реакции Сакагучи как со свободным аргинином, так и с его остатками, содержащимися в пептидах пепсинового гидролизата лизоцима. Расположение опытных точек точно по прямой свидетельствует о том, что в гидролизате, осажденном ТХУ, весь аргинин полностью реагирует с реактивом Сакагучи.
Воздействие Фолина на тирозин не специфично. Кроме тирозина в пептидах с этим реактивом реагируют триптофан, цистеин. Образуемые медью с тетрапептидами и полипептидами биуретовые комплексы облегчают такое взаимодействие. Следовательно, выражение величины активности протеолитических ферментов в тирозиновых эквивалентах и по Фолину, как это иногда делают, неправильно.
Таким образом, мы рассмотрели особенности ферментов как биологических катализаторов, показаны их отличия от небелковых катализаторов, способы измерения активности предложенных ферментов - пепсина и папаина. К сожалению, на данный момент имеется довольно незначительное количество публикаций по исследованию папаина.
1. Березов Т.Т., Коровкин Б.Ф. Биологическая химия: Учебник.- М.: Медицина, 1990.- с.115
2. Основы биохимии: Учебник для студ. биол. спец. ун-тов/под ред. А.А. Анисимова.- М.: Выс.шк., 1986. - с.133-140
3. Фёршт Э. Структура и механизм действия ферментов.- М.: Мир., 1980.- с. 373-388
4. Кочетов Г.А. Практическое руководство по энзимологии.- М., 1989
5. Диксон М., Уэбб Э. Ферменты: пер.англ.- М.: Мир, 1982.- т.1.- с. 370-375
6. Асатиани В.С. Биохимическая фотометрия.- М.: Изд. АН СССР, 1957.- с.248-253
