

 Курсовая работа: Биологическая функция нуклеиновых кислот
Курсовая работа: Биологическая функция нуклеиновых кислот
1.3. Вторичная и третичная структуры ДНК
Растворы ДНК характеризуются аномальной (структурной) вязкостью. В потоке обладают двойным лучепреломлением, что объясняется удлиненной формой молекул ДНК.
В расшифровку структуры ДНК большой вклад внесли исследования Э. Чаргаффа и его сотрудников (1945—1951 гг.). Для разделения оснований, полученных при кислотном гидролизе ДНК, Э. Чаргафф использовал метод хроматографии. Каждое из этих оснований было определено спектрофотометрически. Он впервые определенно заявил, что ДНК обладают выраженной видовой специфичностью. ДНК, выделенные из различных источников, отличаются друг от друга по соотношению входящих в их состав азотистых оснований. Э. Чаргафф сформулировал закономерности состава ДНК, известные под названием правил Чаргаффа. Независимо от происхождения ДНК эти закономерности представляются следующим образом:
1) количество молекул аденина равно количеству молекул тимина (А = Т);
2) количество молекул гуанина равно количеству молекул цитозина (G = С);
3) количество молекул пуриновых оснований равно количеству молекул пиримидиновых оснований (А + G = Т + С);
4) количество оснований с 6-аминогруппами в цепях ДНК равно количеству оснований с 6-гидроксигруппами (А + С = G + Т);
5) отношение (G+С)/(А+Т) резко отличается для разных видов ДНК, но постоянно для клетки одного вида; это отношение называется фактором специфичности.
Фактор специфичности одинаков для ДНК различных органов и тканей одного организма и практически не отличается у разных видов животных и растений в пределах одного класса. У высших растений и животных его величина находится в пределах 0,55—0,93; у бактерий — 0,35—2,73.
Правила Чаргаффа сыграли решающую роль в разработке проблем молекулярной биологии. Именно они легли в основу открытия строения ДНК, ее вторичной структуры.
Прежде чем приступить к рассмотрению этой структуры, необходимо отметить следующее. Интерес к проблеме изучения структуры ДНК возрос в связи с полной неясностью механизма воспроизведения (репликации) ДНК, который отличается очень высокой степенью точности. На основании уже полученных экспериментальных данных предполагалось, что генетическая информация в живой клетке зашифрована (т.е. записана с помощью определенного кода) в специфической последовательности оснований ДНК. Однако было неясно, как воспроизводится такая последовательность.
В 1953 г. Дж. Уотсон и Ф. Крик предложили модель структуры ДНК (рис.2). Она учитывала рентгеноструктурные данные Р. Франклин и М. Уилкинса и "эквимолярность" оснований, открытую Э. Чаргаффом. Модель Уотсона — Крика не только объяснила физико-химические свойства ДНК, но и дала основание высказать предположение о возможном механизме репликации ДНК. Согласно их выводу, молекула ДНК должна быть двухцепочечной. Каждое основание одной цепи "спарено" с лежащим в той же плоскости основанием второй полинуклеотидной цепи. Это спаривание специфично; поскольку количество оснований с гидроксигруппами равно количеству оснований с аминогруппами, то, по Уотсону и Крику, только определенные пары оснований входят в структуру так, что могут образовывать друг с другом водородные связи. Так как аденин содержит аминогруппу, а тимин — гидроксигруппу, цитозин и гуанин — соответственно эти же группы и поскольку они находятся в ДНК в эквимолекулярных количествах, то разрешенными являются только пары А-Т и G-С
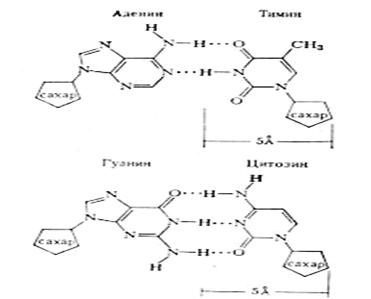
Рис.2. Соединение пар аденина и тимина, цитозина и гуанина в молекуле ДНК при помощи водородных связей
Между А и Т образуются две водородные связи, между G и С - три водородные связи.
Две полинуклеотидные цепи ДНК отличаются одна от другой как последовательностью оснований, так и нуклеотидным составом. Однако основания, стоящие в данном положении в одной цепи, определяют природу основания в другой цепи. Например, если в одной цепи стоит аденин, то напротив него в другой цепи будет располагаться тимин, и наоборот. Если в одной цепи стоит гуанин, то в другой обязательно будет цитозин, и наоборот. Действительно, рентгеноструктурный анализ ДНК показал, что пуриновые и пиримидиновые основания нуклеотидных остатков ДНК лежат в одной плоскости, перпендикулярной продольной оси молекулы, тогда как циклы дезоксирибозы находятся в плоскости, почти перпендикулярной той, в которой лежат циклы оснований.
Явление, при котором последовательность оснований одной цепи однозначно определяет последовательность оснований другой цепи, получило название комплементарности. Таким образом, цепи молекул ДНК комплементарны по отношению друг к другу.
Сформулированный Уотсоном и Криком принцип комплементарности явился универсальным принципом в биологии. Он дал начало развитию новых научных направлений — молекулярной биологии, молекулярной генетике, генной инженерии. Водородные связи обеспечивают способ спаривания оснований, стабильность двухцепочечной системы. Основания плотно упакованы, причем расстояние между центрами оснований, лежащих друг над другом, равно 0,34 нм. На каждый полный виток двойной спирали приходится 10 нуклеотидных пар. Упакованные внутри двойной спирали основания гидрофобны и недоступны молекулам воды. Ионизированные фосфатные группы и гидрофильные остатки дезоксирибозы находятся на поверхности молекулы и контактируют с молекулами воды. Таким образом, двойная спираль стабилизирована не только водородными связями между комплементарными основаниями, но и гидрофобными взаимодействиями между основаниями, расположенными вдоль длинной оси молекулы ДНК. Из-за высокой степени упорядоченности макромолекул ДНК ее иногда называют апериодическим одномерным кристаллом.
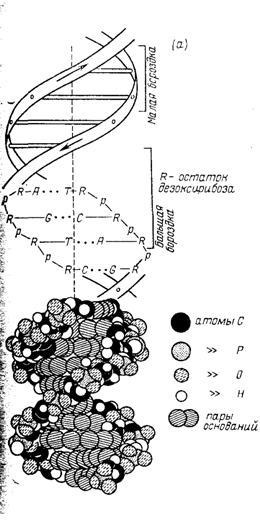
Рис.3 Макромолекулярная структура ДНК
При рентгенографическом исследовании головок сперматозоидов получается такая же дифракционная картина, что и для образцов ДНК, т.е. спираль Уотсона—Крика наблюдается непосредственно в живых клетках.
Модель строения ДНК в настоящее время является общепризнанной. За расшифровку структуры ДНК Дж. Уотсону, Ф. Крику и М. Уилкинсу в 1962 г. была присуждена Нобелевская премия.
Третичная структура ДНК образуется в результате дополнительного скручивания в пространстве двухцепочечной молекулы. Она имеет вид суперспирали или изогнутой (сломанной) двойной спирали.
В настоящее время описаны три формы структуры ДНК: А-, В- и Z-формы (рис.4). Параметры модели Уотсона—Крика соответствуют конформации ДНК в физиологических условиях (В-форме ДНК). Однако при изменении условий среды двойная спираль может принимать другие формы. Так при уменьшении влажности (в препарате образца для рентгеноструктурного анализа) ДНК переходит в А-форму. Этот переход связан с изменением конформации в остатках дезоксирибозы, уменьшением расстояния между фосфатными группами сахарофосфатного остова. Расстояние между парами нуклеотидов вдоль оси спирали, равное 0,34 нм в уотсон-криковской модели, уменьшается (примерно до 0,25 нм при 11 нуклеотидных остатках на один виток спирали). Диаметр спирали увеличивается; изменяются ширина и глубина бороздок; комплементарные пары азотистых оснований образуют с осью спирали угол 20° и, главное, они смещаются к периферии спирали. Поэтому двойная спираль похожа на пологую винтовую лестницу и внутри нее возникает полость диаметром 0,40 нм.
Переход молекулы ДНК из В- в А-форму можно осуществить при понижении активности воды в растворе (при внесении в него органического растворителя, например, этанола). Существует мнение, что В-форма представляет собой некую промежуточную форму двух или большего числа конформаций. Одной из особенностей В-формы, называемой В'-формой, является способность менять в молекуле ДНК положение двух цепей на обратное. Более того, В-форма может существовать в виде как правой, так и левой спирали.
Хотя более стабильными в А- и В-формах являются правозакрученные спирали, существуют довольно устойчивые и левозакрученные спирали ДНК. Одна из таких спиралей была получена в 1979 г. А. Ричем. Из-за нерегулярного зигзагообразного изгиба cахарофосфатного остова она была названа Z-формой (рис.4). Повторяющаяся единица в Z-форме ДНК включает две пары нуклеотидов, а не одну, как в В- и А-формах. Вследствие этого линия, соединяющая фосфатные группы, через каждые две пары нуклеотидов имеет излом и принимает зигзагообразный вид. По сравнению с В-формой в левой Z-форме резко изменен характер стэкинга оснований: сильные и слабые межплоскостные взаимодействия также чередуются. Z-форма может переходить в В-форму при снижении ионной силы раствора, добавлении этанола. Однако вопрос о существовании Z-формы ДНК in vivo и ее биологической роли до конца не выяснен. Высказывается мнение, что переход правозакрученной формы в левозакрученную может служить регуляторным сигналом, контролирующим экспрессию генов.
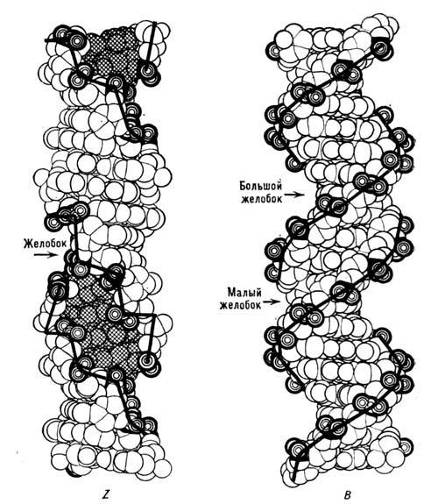
Рис.4. B- и Z-формы структуры ДНК
1.4. Физико-химические свойства ДНК
ДНК — довольно сильная многоосновная кислота, полностью ионизированная
при рН 4,0. Фосфатные группы расположены по периферии. Они прочно связывают
ионы Са![]() и
Мg
и
Мg![]() , амины, гистоны — положительно
заряженные белки. Устойчивость комплементарных пар оснований зависит от
величины рН. Пары оснований наиболее устойчивы в интервале рН 4,0—11,0. За его
пределами двухцепочечная спираль ДНК теряет устойчивость и раскручивается.
, амины, гистоны — положительно
заряженные белки. Устойчивость комплементарных пар оснований зависит от
величины рН. Пары оснований наиболее устойчивы в интервале рН 4,0—11,0. За его
пределами двухцепочечная спираль ДНК теряет устойчивость и раскручивается.
Молекулярная масса ДНК неодинакова и зависит от источника ее
получения. К тому же даже при самых тщательных и щадящих процедурах выделения
ДНК подвергается некоторой деградации. Препараты, полученные современными
методами из тканей животных и растений, имеют молекулярную массу 6 10![]() —10 10
—10 10![]() . Однако истинная
молекулярная масса ДНК животных и растений, определенная по вязкости и по длине
молекул, значительно выше и достигает десятков миллиардов.
. Однако истинная
молекулярная масса ДНК животных и растений, определенная по вязкости и по длине
молекул, значительно выше и достигает десятков миллиардов.
