Раствор
С: смешивали по 200 мл растворов А и В, перемешивали в пластиковой емкости на
магнитной мешалке в течение 30 мин., фильтровали полученный раствор С через
плотный бумажный фильтр в пластиковую посуду. Раствор С использовали для
анализа. Хранили в холодильнике, в течение надели. Перед каждым употреблением
фильтровали через бумажный фильтр.
3. Обсуждение
результатов
3.1 Отбор
штаммов микроорганизмов-деструкторов
фосфорорганических
соединений
Для
скрининга использовали коллекцию штаммов Pseudomonas и Alcaligenes лаборатории института биохимии и
физиологии растений и микроорганизмов РАН. В качестве субстрата использовали
МФК.
Образование
зон «просветления» на среде, содержащей МФК, представлено на рисунке 6.
Рисунок
6 – Образование зон «просветления» различными штаммами
Как
видно из рисунка 6, наибольшей способностью к усилению растворения МФК обладают
штаммы 2-79, Pf-5, P54, 1C7,
относящиеся к роду Pseudomonas fluorescens. Наилучшая способность к растворению
метилфосфонатов отмечается у штамма Sm11 , относящегося к роду Alcaligenes sp.
Таким
образом, биодоступность МФК можно достичь внесением в почву штамма Sm11 бактерий рода Alcaligenes sp. Поэтому для изучения эффективности биодеструкции
фосфорорганических соединений выбран именно этот штамм.
3.2 Биодеструкция
метилфосфоновой кислоты и её кислых эфиров
Из
литературных данных известно, что неорганический фосфат ингибирует потребление
органофосфонатов в качестве источника фосфора разложение МФК [33-35]. При
выращивании посевного материала на агаризованной среде, по-видимому, возможна
экстракция из агара и накопление в клетках более доступных, чем МФК, источников
фосфора, которые могут попадать в экспериментальную среду вместе с посевным
материалом. Более доступный фосфор может находиться на поверхности агара,
образуясь в процессе жизнедеятельности культуры из МФК при наращивании
биомассы. Поэтому, к тому времени, как биомасса на агаризованной среде нарастет
до необходимого уровня, в клетках посевного материала могут быть запасные
источники фосфора, за счет которых культура будет расти в среде с МФК, не
разлагая его. Чтобы исключить все эти факторы необходимо иметь клетки посевного
материала, голодные по фосфору. Для решения этой проблемы необходимо суспензию
клеток, используемую в качестве посевного материала, инкубировать в среде с
глутаматом (источником углерода), но без источника фосфора в течение трех
суток. В это время можно считать клетки голодными по фосфору.
Подготовленный посевной материал использовали для
засева опытных колб с концентрацией МФК (метилфосфонатов) 0,3 г/л. Рост культур
контролировали по изменению оптической плотности и контролировали изменение
содержания органического фосфора и значение рН. По данным измерений оптической
плотности и концентрации загрязнителя строили кривые роста культур и
потребления источника фосфора (рисунок 7 и 8).
ГЛУТАМАТ
контроль
80
20
60
время, ч
40
0
2
3
5
4
1
оптическая плотность, отн.ед.
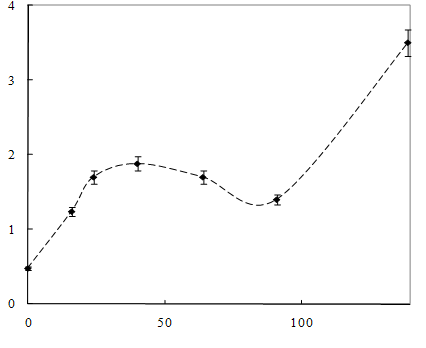
Рисунок
7 – Динамика роста штамма Sm11 в среде, содержащей МФК
концентра-
ция,
мг/л
время, ч
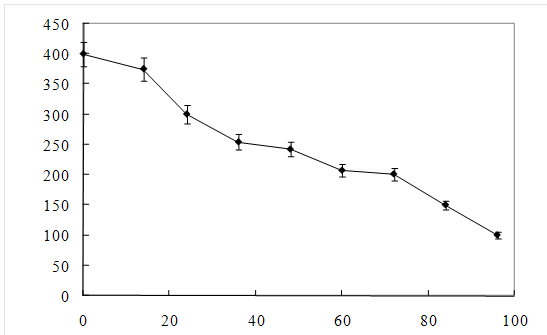
Рисунок 8 – Изменение концентрации МФК
В рассматриваемых системах различают следующие фазы
размножения:
лаг-фаза – период между засевом и началом
размножения;
экспоненциальная фаза – период размножения
микроорганизмов с постоянной скоростью;
фаза гибели – период, в течение которого популяция
погибает.
Как видно из рисунка 7, рост культуры на среде,
содержащей МФК, начинался после непродолжительной лаг-фазы – не более 10 часов,
затем наблюдался значительный рост микроорганизмов. Концентрация МФК уменьшается
в 2 раза в течение 40 -45 часов (рисунок 8). После 58 – 62 часов рост культуры
и потребление МФК прекращалось, что обусловлено недостатком в питательной среде
источника углерода. При добавлении глутамата натрия рост клеток возобновлялся,
и наблюдалось дальнейшее уменьшение концентрации МФК.
Для оценки возможности
биодеструкции изопропилового, изобутилового и пинаколилового эфиров
использовали отобранный ранее штамм Sm11 рода Alcaligenes sp.
Начальная концентрация кислого эфира составляла 0,2 г/л.
Рост культур контролировали по изменению оптической плотности. По данным
измерений оптической плотности и концентрации загрязнителя строили кривые роста
культур и потребления кислых эфиров.
Динамика роста культуры и потребления изопропилового
эфира представлена на рисунке 9 и 10.


 Дипломная работа: Разработка методов и средств реабилитации объектов отравляющих веществ
Дипломная работа: Разработка методов и средств реабилитации объектов отравляющих веществ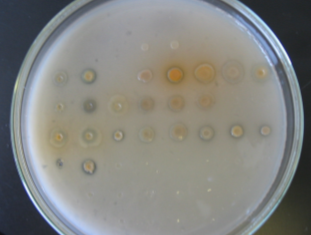
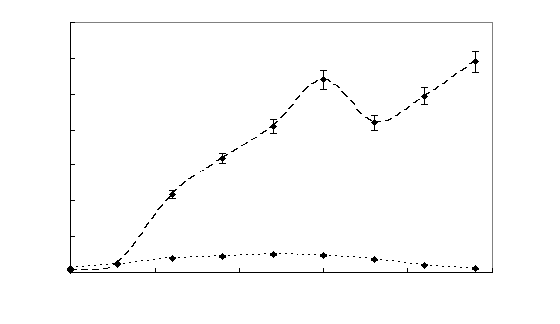
 Рисунок
7 – Динамика роста штамма Sm11 в среде, содержащей МФК
Рисунок
7 – Динамика роста штамма Sm11 в среде, содержащей МФК